فرگشت
فرگشت یا تکامل در حوزهٔ زیستشناسی بهمعنی دگرگونی در یک یا چند ویژگی فنوتیپ و وراثت است که طی زمان در جمعیتهای موجودات زنده رخ میدهد.[1] این ویژگیهای فنوتیپی، که از نسلی به نسل بعد جابجا میشوند؛ صفات کالبدشناسی (آناتومی)، بیوشیمیایی و رفتار را تعیین میکنند. وقوع تکامل منوط به وجود بستری از گوناگونی ژنی در جمعیت است. این بستر ممکن است از جمعیتهای دیگر تأمین شود؛ که به شارش ژن شهرت دارد.[2][3][4][5] همچنین ممکن است گوناگونی ژنی از درون جمعیت، با سازوکارهایی چون جهش یا نوترکیبی پدید آید. با توجه به اینکه ویژگیهای فنوتیپی متفاوت، احتمال بقا و تولیدمثل را تحت تأثیرات متفاوتی قرار میدهند؛ انتخاب طبیعی میتواند سبب فراگیری ژنوتیپهای نو در خزانهٔ ژنی شود؛ و چهرهٔ فنوتیپی جمعیت را به تدریج تغییر دهد.[1] انتخاب جنسی ممکن است به همهگیری ژنهایی ختم شود که نقش مثبتی در افزایش بقای جاندار نداشته باشند؛ و دیگر سازوکارهای تکاملی همچون همبستگی ژنی و رانش ژن، ممکن است ژنهایی را انتخاب کنند که امتیاز مستقیمی برای بقا یا تولیدمثل جاندار ارائه نمیدهند.[6][7]
| بخشی از مجموعه مقالههای |
| زیستشناسی فرگشتی |
|---|
 |
|
درگاه.ویکیپروژه |
رویداد فرگشت بدان معناست که تمام موجودات زنده با همهٔ تنوعی که دارند، از نیاکانی مشترک پدیدار گشتهاند.[8] فرگشت علت پدیدهٔ گونهزایی است؛ که طی آن یک گونهٔ اجدادی منفرد به دو یا چند گونهٔ متفاوت تقسیم میشود. گونهزایی در شباهتهای ساختاری، جنینی و ژنتیکی جانداران؛ پراکندگی جغرافیایی گونههای مرتبط با هم، و ثبت سنگوارهای تغییرات، قابل مشاهدهاست. نیای مشترک جانداران امروزی تا بیش از ۳٫۵ میلیارد سال پیش؛ یعنی از زمان پیدایش حیات روی زمین، قدمت دارد.[9][10][11][12] تکامل چه به صورت درونجمعیتی و چه به صورت گونهزایی میان جمعیتها، از طرق گوناگونی روی میدهد؛ آهسته و پیوسته به نام انتخاب انباشتی، یا به سرعت از یک موضع ایستا تا موضع بعدی؛ که تعادل نقطهای خوانده میشود.
مطالعهٔ علمی فرگشت (تکامل) از نیمهٔ سدهٔ نوزدهم آغاز شد، زمانی که پژوهشها روی ثبت سنگوارهای و گوناگونی جانداران، بسیاری از دانشمندان را متقاعد کرد که میبایست گونهها به نحوی تکامل یابند. بر مبنای سنگوارهها مییابیم که جانداران امروزی متفاوت از گذشته هستند و به میزانی که به گذشتههای دورتر مینگریم، فسیلها متفاوتتر میشوند.[13][14] سازوکارهای پیشبرندهٔ تکامل همچنان نامشخص باقیماندند؛ تا سال ۱۸۵۸ که نظریهٔ انتخاب طبیعی، بهطور مستقل توسط چارلز داروین و آلفرد راسل والاس ارائه شد. در اوایل قرن بیستم، تئوریهای داروینی تکامل با ژنتیک، دیرینشناسی و سامانهشناسی ادغام شدند که با پیوستن یافتههای بعدی چون زیستشناسی مولکولی؛ تحت عنوان تلفیق تکاملی جدید به اوج رسید.[15] این تلفیق به یک بنیان اصلی در زیستشناسی بدل شد؛ چنانکه تبیینی منسجم و یکپارچه، برای تاریخ و تنوع زیستی حیات روی زمین، فراهم ساخت.[16][17][18]
امروزه تکامل در شاخههای مختلف علوم زیستی چون زیستشناسی بقا، جنینشناسی، بومشناسی، فیزیولوژی، دیرینشناسی و پزشکی مطالعه و بهکاربسته میشود. به علاوه، تکامل بر دیگر حیطههای مطالعات بشری، همچون کشاورزی، انسانشناسی، فلسفه و روانشناسی نیز اثرگذار بودهاست.
نظریهٔ فرگشت پارادایم حاکم بر زیستشناسی نوین است که پایه و اساس آن را تشکیل میدهد و با شواهد بسیاری پشتیبانی میگردد؛ بهطوریکه به گفته دبژنسکی «هیچ چیز در زیستشناسی جز در پرتو آن معنا نمییابد».[19] همزمان در جزئیات اختلاف نظر هست و پرسشهای بسیاری نیازمند پاسخ هستند که زیستشناسان تکاملی به آنها میپردازند. زیستشناسان تکاملی واقعیت وقوع تکامل را مستند کرده، و همچنین نظریاتی در شرح علل آن توسعه داده و میآزمایند.
واژهشناسی
واژهٔ عربی تکامل به معنای «کاملشدن» است. لغتنامهٔ دهخدا معنی آن را «تمامشدن» ذکر میکند.[20] این واژه نخستین بار توسط مترجمان دورهٔ قاجار به عنوان برابر فارسی «evolution» در زبانهای اروپایی بهکار رفت.
این در حالیست که واژهٔ «evolution» بهمعنای «برآمدن» و «تحوّل» است و اشاره به وجودآمدن چیزی از چیز دیگر دارد، مثل وجودآمدن بخار از آب. «Evolution»، حاکی از «کمالیافتن» جانداران نیست. این واژه هیچ بار اخلاقی ندارد بلکه تنها تغییر جانداران برای انطباق و سازگاری بیشتر با محیط زیست خود را نشان میدهد، زیرا در یک محیط، ویژگیهای خاصی مبنای تطابق و سازگاری محسوب میشوند و در محیط دیگر ویژگیهای دیگری.
لذا فرگشت همتراز مناسبتری برای Evolution است.[21]
«فرگشت» از ساختههای فرهنگستان دوم است.[22] این واژه در آثار نویسندگانی همچون داریوش آشوری بهکار برده شدهاست.[23] بعدها برابرهای دیگری نیز از سوی پژوهشگران پارسیزبان برای Evolution پیشنهاد شد؛ از آن جمله واژه «برآیش» که توسط ابراهیم هرندی، روانزیستشناس، پیشنهاد شد.[24]
در منابع و رسانههای فارسیزبان بهجای «تکامل» یا «فرگشت» از واژههایی همچون «ترقیکردن» و «ارتقاءیافتن» نیز استفاده شدهاست، در بعضی برنامههای علمی پخششده توسط صدا و سیمای جمهوری اسلامی ایران از عبارت «سازگاری و تغییر» نیز استفاده شدهاست.[25]
در زبان اردو و زبان سندی که از زبانهای همریشه و همخانواده با زبان فارسی هستند، به این نظریه «ارتقا» گفته و نوشته میشود.[26][27]
تاریخ اندیشهٔ تکاملی
آرای قدما
این پیشنهاد که یک نوع جانور ممکن است از جانوری از نوع دیگر منشأ گرفته باشد، در آرای برخی فلاسفهٔ پیشاسقراطی چون آناکسیماندروس و امپدوکلس به چشم میخورد. در تضاد با این دیدگاههای مادهگرایانه، ارسطو همهٔ اشیاء طبیعی و نه تنها موجودات زنده را، فعلهایی ناقص از حالات طبیعی ممکن میدانست؛ که ثابت بودند. او این حالات ثابت را، صورت یا مثال؛ یا (در ترجمهٔ لاتین) گونه مینامید. این دیدگاه بخشی از فهم الهی ارسطو از طبیعت بود، که در آن همهٔ اشیاء نقشی غایتمند در نظام الوهیت جهان ایفا میکردند. البته وی ادعا نکرد که هر جانور، با یک صورت مابعدالطبیعی خاص تناظر یکبهیک دارد؛ بلکه نمونههایی ذکر کرد که چگونه انواع جدیدی از جانداران میتوانستهاند به وجود آیند. تعابیر گوناگون از نظریهٔ ارسطو، پس از ترکیب با تعالیم مسیحی، فهم متعارف در قرون وسطا شد.
سدههای هفدهم و هجدهم
در قرن هفدهم، روش جدید علم نوین، رویکرد ارسطویی به طبیعت را رد کرد، و در جستجوی تشریح پدیدههای طبیعی در قالب قوانین طبیعت برآمد؛ که برای همهٔ محسوسات یکسان، بینیاز از فرض هیچگونه ردهٔ ثابت طبیعی، یا نظام الهی جهان بودند. امّا این رویکرد جدید به کندی در زیستشناسی ریشه دواند؛ که به آخرین سنگر برای درک ثابت از اشیای طبیعی مبدّل گشت.
جان ری اصطلاح گونه را که آن زمان متروک شده بود، برای اطلاق به انواع ثابت جانوری و گیاهی بهکار برد؛ اما برخلاف ارسطو، او اکیداً هر نوع از موجودات زنده را به عنوان یک گونه، معرفی کرد. ری پیشنهاد کرد که هر گونه را میتوان به مدد ویژگیهایی شناخت که در هر نسل، هنوز باقی هستند و خود را حفظ میکنند. این گونهها توسط خدا طراحی شده بودند؛ اما تفاوتهایی نشان میدادند که بر اثر شرایط محلی پدیدار شده بود. در طبقهبندی زیستی جانداران توسط کارل لینه که در ۱۷۳۵ معرفی شد نیز؛ گونهها ثابت بر اساس طرح الهی در نظر گرفته شدند.
برخی دیگر از طبیعتگرایان این زمان، به گمانهزنی دربارهٔ تغییرات تکاملی گونهها بر اساس قوانین طبیعت پرداختند. در ۱۷۵۱، موپرتوئی از تغییراتی که حین تولیدمثل رخ میدهند نوشت؛ و انباشت این تغییرات را طی نسلها نیروی پیشران تکامل دانست. بوفون پیشنهاد کرد که ممکن است گونهها بتوانند به جاندارانی متعدد تحلیل یابند؛ و اراسموس داروین این فرضیه را مطرح کرد که ممکن است تمام جانوران خونگرم از یک میکروب منفرد (یا به بیان او از یک فیلامنت) مشتق شده باشند.
نظریه تکاملی لامارک:استحاله ماهوی
نخستین طرح تکاملی پخته، نظریهٔ «ترانسموتاسیون» یا «استحاله ماهوی» ژان لامارک بود که به سال ۱۸۰۹ در کتاب فلسفه جانورشناسی منتشر شد. لامارک تجسم کرده بود که همچنانکه نسل خودانگیخته به تولید اشکال سادهٔ حیات ادامه میدهد؛ پیچیدگی بیشتر در دودمانهای موازی با یک تمایل پیشروندهٔ موروثی توسعه مییابد. بهطوریکه تغییرات موروثی بر اثر استفاده یا عدم استفادهٔ والدین، دودمانها را به سازش بیشتر با محیط میکشاند.[28][29] این فرایند پیشنهادی، بعدها لامارکیسم خوانده شد.[28][30][31][32] اردوگاه طبیعتگرایان این آراء را به علت فقدان شواهد تجربی، محکوم میکردند. کوویه تأکید کرد که گونهها غیرمرتبط و ثابت هستند؛ و شباهتهای آنها نشانگر طرح الهی و بر اساس نیازهای عملکردی آنهاست.
در همین زمان، ایدهٔ طراح نیکخواه جان ری؛ توسط ویلیام پِیلی در جهت الهیات طبیعی پرورانده شد. پیلی استدلال کرده بود که سازشهای پیچیدهٔ بدن جانداران، نشانههای طرح الهی هستند. چارلز داروین جوان، این آرای پیلی را تحسین کرد.[33][34][35]

نظریه تکاملی داروین:انتخاب طبیعی
فرضیهٔ تکاملی چارلز داروین، نقطهٔ عطفی در مسئلهٔ ثبوت یا تکامل انواع بهشمار میرود. داروین متأثر از نظرات توماس مالتوس در رسالهٔ اصل جمعیت؛ به این نتیجه رسید که رشد جمعیت به آسانی میتواند به «تنازع بقا» منجر شود؛ و در این رقابت زادههای با مطلوبیت بیشتر، بر دیگران برتری خواهند داشت. محدودیت منابع باعث میشود تا در هر نسل بسیاری از زادهها پیش از رسیدن به سن تولیدمثل تلف شوند. این ایده میتوانست گوناگونی جانوران و گیاهان را، در عین اشتقاق از اجداد مشترک توضیح دهد؛ و در عین حال این شرح تنها با استفاده از قوانین طبیعی که برای همهٔ اشیاء یکسان بودند، فراهم میشد.[37][38][39][40]
داروین در ۱۸۳۸ به نظریه انتخاب طبیعی دست یافت، امّا آن را منتشر نکرد؛ تا اینکه در ۱۸۵۸ نامهای از یک زیستشناس جوان به نام آلفرد والاس به دستش رسید که در آن نظریه مشابهی شرح داده شده بود. والاس در این نامه به منظور انتشار فرضیهٔ خود از داروین تقاضای کمک کرده بود. آنها دو مقالهٔ جداگانه برای قرائت در یکی از گردهمآییهای علمی انجمن لینهایهای لندن ارسال کردند.[41]
داروین و والاس هیچکدام در این گردهمایی شرکت نکردند و در آن زمان تعداد اندکی از آن دو مقاله مطلع شدند. سرانجام در اواخر سال ۱۸۵۹، کتاب داروین به نام «دربارهٔ خاستگاه گونهها از طریق انتخاب طبیعی» منتشر شد و به تفضیل به شرح تکامل داروینی پرداخت. توماس هاکسلی، آناتومی مقایسهای و دیرینشناسی را به کار گرفت تا نشان دهد که انسان و کپیها نیاکان مشترکی دارند. این نتیجهگیری تلویحی که انسان جایگاهی یگانه و خاص در جهان ندارد، بسیاری را برآشفت.[42]
تولد و رشد دانش ژنتیک
سازوکارهای دقیق وراثت، و منشأ خصائص جدید همچنان معما باقیماندند. برای حل این معما، داروین تئوری پانژنز را مطرح کرد.[43] در ۱۸۶۵، گرگور مندل گزارش کرد که ویژگیهای فنوتیپی با یک الگوی پیشبینیپذیر ناشی از تفکیک و جورشدن مستقل تعدادی از عناصر به ارث میرسند. (این عناصر بعدها ژن خوانده شدند) قوانین وراثت مندلی در نهایت تئوری پانژنز داروین را رد کرد.[44] آگوست وایزمن تمایز مهمی میان سلولهای ژرمینال (اسپرم و تخمک) و سوماتیک (پیکری) بدن قائل شد؛ و مشخص ساخت که وراثت تنها از طریق خط ژرمینال عمل میکند.
هوگو دووری تئوری پانژنز داروین را با تمایز سوماژرمینال وایزمن، مربوط کرد و این فرضیه را مطرح ساخت که اثر پانژنها که در هستهٔ سلول متمرکز هستند، با بیان شدن به سیتوپلاسم منتقل میشود و ساختار سلول را تغییر میدهد. دووری همچنین معتقد بود که انتقال صفات متنوع زادهها طی خط ژرمینال، از طریق وراثت مندلی انجام میپذیرد.[45] در توضیح منشأ خصائص جدید، دووری فرضیهٔ جهش را توسعه داد، که منجر به ایجاد اختلافی موقتی میان دانشمندان قائل به تکامل داروینی، و زیستسنجشان پیروی دووری شد.[29][46][47]
در آغاز قرن بیستم، پیشگامان ژنتیک جمعیت چون جان هالدین، سؤال رایت و رونالد فیشر مبنایی مستحکم و محاسباتی برای تکامل فراهم ساختند؛ و تضاد ظاهری میان نظریهٔ داروین، جهشهای ژنتیکی و وراثت مندلی از میان رفت.[48]
تلفیق تکاملی جدید
در دهههای ۱۹۲۰ و ۱۹۳۰، تلفیق تکاملی جدید، مفاهیم انتخاب طبیعی، جهش و وراثت مندلی را به هم مرتبط ساخت و به یک نظریهٔ یکپارچه تبدیل شد که بهطور کلی در تمام شاخههای علوم زیستی به کار رود. تلفیق تکاملی جدید قادر به توضیح الگوهای مشاهدهشده در گونهها و جمعیتها، تغییرات فسیلی در دیرینشناسی، و حتی سازوکارهای پیچیدهٔ سلولی در جنینشناسی بود.[29][49]
پیشرفتهای متاخر
در ۱۹۵۳ جیمز واتسون و فرانسیس کریک با کشف ساختار مولکول DNA مبنایی دقیق برای تبیین فعالیتهای آن فراهم ساختند.[50] زیستشناسی مولکولی فهم ما را از روابط بین ژنوتیپ و فنوتیپ بهبود بخشید. پیشرفتها در فیلوژنتیک و سامانهشناسی، از طریق انتشار و بهکارگیری درختهای تکاملی، تحول صفات را در یک چهارچوب مقایسهای و آزمودنی ترسیم کرد.[51][52] در ۱۹۷۳،تئودوزیوس دابژانسکی زیستشناس تکاملی نوشت: «هیچچیز در زیستشناسی معنا پیدا نمیکند، مگر در پرتو تکامل»، زیرا تکامل چیزهایی را که زمانی نامرتبط با هم به نظر میرسیدند؛ در قالب یک شرح منسجم جای میدهد که قادر به توصیف و پیشبینی حقایق مشاهدهشده در خصوص حیات است.[53]
از آن زمان، تلفیق تکاملی جدید گسترش یافته تا در مقیاس سازماندهی زیستی، از ژن تا گونه را توضیح دهد. این گسترش اِکو-اِوو-دِوو نامیده شدهاست.[15][15][54][55]
انتقادات واردشده بر فرگشت
وراثت
- برای اطلاعات زمینهای و مطالب مرتبط با این بخش مقاله ژنتیک را ببینید.
ژنتیک مندلی
بر اساس پژوهشهای مِندل که در ۱۸۶۶ انتشار یافت، هر فرد جاندار برای هر صفت خود دو حالت(الل) دارد که یکی از آنها را از پدر و دیگری را از مادر دریافت کردهاست. اصطلاح ژنوتیپ اشاره به نوع اللهای هر فرد،[56] و فنوتیپ اشاره به شکل بروز آن اللها دارد.[57] مندل افراد خالص را که دو الل یکسان داشتند هُموزایگوت، و افرادی را که دو الل متفاوت داشتند هِتِروزایگوت مینامید. هموزایگوتها ناشی از خودلقاحی بودند و P نامیده میشدند. دودمانهای هتروزایگوت F1 و F2 نیز به ترتیب از دگرلقاحی و خودلقاحی نسل پیشین خود حاصل میشدند. مندل اللی را که در F1 اثر خود را بهطور کامل نشان میداد غالب، و اللی را که تنها در F2 ظاهر میشد مغلوب نام نهاد. بر اساس این تحقیقات قوانین سهگانه مشهور به قوانین مندل یا قوانین وراثت پایهریزی شدند: قانون اوّل یا قانون یکنواختی، قانون دوم یا تفکیک، و قانون سوم یا جورشدن مستقل.
اساس سلولی وراثت
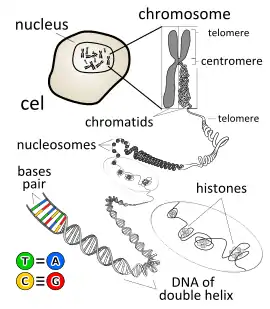
ساختار کروموزوم بر اساس «مدل گردنبند مروارید» توصیف میشود، که نخ را DNA و نوکلئوزومها (دانهها) ی آن را DNA احاطه شده به وسیله پروتئین تشکیل میدهد. اساس سایتوژنتیکی وراثت بر اساس فرضیه سال ۱۹۰۲ ساتون و بوواری مبنی بر اینکه کروموزومها، ماده ژنتیکی را طی لقاح حمل میکنند، شکل گرفت؛ با این وجود اساس مولکولی وراثت همچنان نامشخص باقیماند، چرا که ماهیت بیوشیمیایی ماده ژنتیکی مشخص نشده بود.
اساس مولکولی وراثت
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، DNA و ژن را ببینید.
آزمایشهای اِیوری به درک اساس مولکولی وراثت کمک شایانی کرد. او در ۱۹۴۴ دریافت که ماده ژنتیکی نوعی نوکلئیک اسید به نام DNA است. ساختار فضایی مولکول DNA در ۱۹۵۳ توسط واتسون و کریک تشریح شد. بر اساس «مدل مارپیچ دوگانه» واتسون و کریک، مولکول DNA همچون نردبانی مارپیچ است که هر پله آن را یک جفت باز، و دو نرده آن را توالیهای قند-فسفات تشکیل میدهد. باز مذکور یک پورین(آدنین A یا گوانین G)؛ یا پیریمیدین(سایتوزین C یا تیمین T) است. قند مذکور نیز پنتوزی به نام دئوکسیرایبوز است. به منظور همانندسازی DNA دو رشته این مولکول میبایست توسط آنزیم هلیکاز از هم باز شوند.[56] بر این اساس ژن به صورت تکرشتهای که به عنوان الگو برای همانندسازی قرار میگیرد، بررسی میشود. ژن متشکل از توالیهای اگزون و اینترون است که همچون واگنهای قطار دنبال هم سوار شدهاند. از آنجا که مولکولهای قند و فسفات در رمزگذاری صفات نقشی ندارند، یک ژن را میتوان تنها توسط بازهایش به صورتهایی چون…AUGCCTA نمایش داد.
نقش وراثت در تکامل صفات
مطابق با نظریه ۱۹۴۰ بیدل و تِیتوم تحت عنوان «نظریه یک ژن-یک آنزیم»، هر ژن رمز ساخت یک آنزیم را داراست. این نظریه امروزه به صورت «نظریه یک ژن-یک پلیپپتید»، تصحیح شدهاست. نتیجه تکاملی توارث آناست که ژنها رمز ساخت پروتئینها را حمل میکنند، و پروتئینها مسئول واکنشهای جنینی و پس از تولد مؤثر در صفات ساختاری، فیزیولوژیک، بیوشیمیایی و رفتاری (در مورد سلولهای مغزی) جانداران هستند. این صفات شانس بقا و تولیدمثل را به درجات مختلفی تحت تأثیر قرار میدهند؛ و در نتیجه فراوانی اللها طی نسلهای متمادی تغییر میکند.[58][59]
نظریه تکاملی لامارک برای مدتها به علت مردود شمرده شدن وراثت صفات اکتسابی مطرود بود؛ با وجود این یافتههای چند دهه اخیر نشان دادهاند که برخی صفات اکتسابی میتوانند بدون تغییر در توالی نوکلئوتیدهای DNA به ارث برسند. سازوکارهای این نوع توارث از قبیل متیلاسیون DNA، خاموش شدن ژن از طریق تداخل RNA، و ساختمان سوم پروتئینی در پریونها[60][61]؛ همگی تحت عنوان توارث اپیژنتیک طبقهبندی میشوند و در تکامل نقش دارند.[62] همچنین به جز توارث اپیژنتیک؛ اقسام توارثی دیگری نیز در تکامل شرکت میکنند که مستقیماً تحت کنترل ژنها نیستند و از جمله آنها میتوان به DIT و سیمبیوژنز اشاره نمود.[63][64]
گوناگونی ژنی


فنوتیپ در هر فرد از ژنوتیپ و اثرات محیط خارج او، منشأ میگیرد. بخش عمدهای از گوناگونیها در فنوتیپ به دلیل تفاوتها در ژنوتیپها است. فرگشت به معنای تغییرات در گوناگونیهای ژنتیکی میباشد. در طول زمان ممکن است فراوانی هر الل (ژن) نسبت به بقیه اللهای مربوطه، کم یا زیاد شود.[65] هنگامیکه یک الل به تثبیت یا ثبات وراثتی (به انگلیسی: fixation) برسد، میزان گوناگونی الل، صفر خواهد شد. انتخاب طبیعی زمانی موجب فرگشت میشود که گوناگونیهای ژنتیکی کافی در جمعیت وجود داشته باشد.[66] اصل هاردی- وینبرگ (به انگلیسی: Hardy-Weinberg principle)، چگونگی باقی ماندن گوناگونیها در جمعیت با وراثت مندلی را نشان میدهد. فراوانی اللها در غیاب انتخاب، جهش (موتاسیون)، مهاجرت و رانش ژنتیکی ثابت باقی میماند.[67]
جهش
جهش به معنای تغییر در توالیهای DNA در سلول است. جهش ممکن است هیچ اثری نداشته باشد، محصول ژنی را تغییر دهد یا عملکرد ژن را مختل سازد. مطالعات بر روی مگس سرکه نشان میدهد که اگر جهش موجب تغییر محصول ژن (پروتئین) شود، این مسئله خطرناک خواهد بود زیرا ۷۰٪ این جهشها اثرات تخریبی دارند.[68]
جهشها میتوانند موجب مضاعف شدند بخشی ار کروموزوم (نوترکیبی ژنتیکی) شوند که در نتیجه، کپیهایی اضافی از ژن وارد ژنوم خواهد شد.[69] همین کپیهای اضافی از ژن، منشأ مناسبی برای ایجاد ژنهای جدید از راه فرگشت هستند. بسیاری از خانوادههای ژنی (به انگلیسی: gene families)به همین ترتیب از ژنهای قدیمیتر (اجدادی) به وجود آمدهاند.[70] به عنوان مثال، چشم انسان از محصول ۴ ژن برای دیدن استفاده میکند. محصول ۳ ژن در دیدن رنگ و محصول یک ژن در دیدن در نورکم (شب) نقش دارد. همگی این ژنها از یک ژن اجدادی به وجود آمدهاند.[71] ژنهای جدید از راه مضاعف شدن ژن اجدادی و جهش در ژن تازه به وجود آمده (مضاعف شده) به وجود میآیند.
جنسیت و نوترکیبی
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، تکامل تولیدمثل جنسی و نوترکیبی ژنتیکی را ببینید.
در جاندارانی که تکثیر غیر جنسی دارند، ژنها با یکدیگر به ارث میرسند و نمیتوانند با ژنهای سایر جانداران در هنگام تولید مثل، مخلوط شوند اما در جاندارانی که تکثیر جنسی دارند، کروموزومهای والدی (مادری و پدری) با یکدیگر مخلوط میشوند. در فرایندی به نام نوترکیبی هومولوگ (به انگلیسی: homologous recombination)، این جانداران، DNA را بین کروموزومهای جفت شده (به انگلیسی: matching chromosomes) مبادله میکنند.[72] پس جنسیت موجب افزایش گوناگونی ژنتیکی و افزایش سرعت فرگشت (تکامل) میشود.[73]
شارش ژنی
شارش ژنی (به انگلیسی: Gene flow) به معنای تبادل ژن بین جمعیتها و گونهها میباشد.[74] بنابراین به عنوان منبعی برای گوناگونیها است. شارش ژنی میتواند بر اثر مهاجرت جمعیتها از جایی به جای دیگر اتفاق بیفتد. انتقال ژن بین گونهها شامل تشکیل جانداران دورگه (هیبرید) و انتقال افقی ژنها (به انگلیسی: Horizontal gene transfer) است. انتقال افقی ژنها به معنای انتقال ژن از یک جاندار به جاندار دیگر است بهطوریکه جاندار جدید زاده یا فرزند جاندار دیگر نباشد. انتقال افقی ژنها، بیشتر در باکتریها دیده شدهاست. انتقال ژنهای مقاومت به آنتیبیوتیکها از این راه اتفاق میافتد.[75] از این رو، در پزکش اهمیت فراوانی دارد. در برخی از یوکاریوتها مانند ساکارومایسس سرویزیه (به لاتین: Saccharomyces cerevisiae) نیز انتقال افقی ژنها اتفاق میافتد. ویروسها هم در انتقال DNA از یک جاندار به جاندار دیگر نقش دارند.[76] اکتساب کلروپلاست (سبزینه) و میتوکندری توسط سلولهای یوکاریوتی، نمونهای از جذب مقدار زیاد ژن است. میتوکندری و کلروپلاست منشأ باکتریایی دارند. ممکن است یوکاریوتها بر اساس انتقال افقی ژنها بین باستانیان (آرکئا) و باکتریها به وجود آمده باشند.[77]
سازوکارها
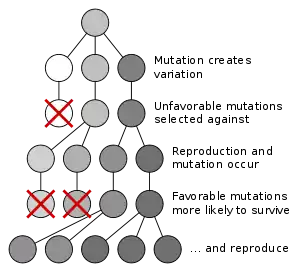
از دیدگاه نوداروینی، تکامل زمانی اتفاق میافتد که فراوانی اللها درون یک جمعیت از نژادهای مختلف-که بتوانند با یکدیگر آمیزش کنند- تغیر کند.[67] به عنوان مثال، الل رنگ سیاه در جمعیتی از پروانهها شایع تر میشود. برخی از مکانیزمهایی که میتوانند منجر به تغییر در فرکانس اللها شود عبارتند از: انتخاب طبیعی، رانش ژن، شارش ژن، سفر رایگان ژنتیکی و تورش ناشی از جهش.
انتخاب طبیعی
تکامل به وسیله انتخاب طبیعی فرایندی است که طی آن جهشهای ژنتیکی بهبود دهنده بقا و تولیدمثل، در جمعیت شایع تر میشوند؛ و در نسلهای موفق یک جمعیت باقی میمانند. این سازوکار اغلب «بدیهی» خوانده میشود؛ زیرا ضرورتاً از سه واقعیت ساده زیر پیروی میکند:
- گوناگونیهای ارثی درون هر جمعیت از جانداران وجود دارند.
- جانداران بیش از تعدادی که بتوانند زنده بمانند، فرزند تولید میکنند.
- این فرزندان در توانایی زنده ماندن و تولیدمثل تفاوت دارند.
چنین وضعی منجر به شکلگیری رقابت بین جانداران برای زنده ماندن و تولیدمثل میشود؛ در نتیجه جاندارانیی که خصائص ارثی شان به آنها مزیتی نسبت به رقبا ارائه میدهد، این خصائص مفید را به نسل بعد منتقل میکنند؛ در حالی که خصائص غیرمفید به نسل بعد نمیرسند.[78]
مفهوم مرکزی در انتخاب طبیعی، مفهوم سازواری جاندار است.[79] سازواری با قابلیت بقا و تولید مثل جاندار اندازه گرفته میشود؛ که میزان مشارکت آن را در نسل بعد تعیین میکند.[79] سازواری نه با تعداد کلی زادهها، بلکه با نسبتی از نسلهای بعدی که ژنهای جاندار را حمل میکنند؛ تعیین میشود.[80] برای مثال جانداری که به خوبی زنده بماند و به سرعت تولید مثل کند، اما زادههایی کوچک و ضعیف بر جا گذارد؛ مشارکت ژنتیکی ناچیزی در نسلهای بعدی خواهد داشت، و در نتیجه سازواری اش پایین در نظر گرفته میشود.[79]
رانش ژن
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، رانش ژن و اندازه جمعیت مؤثر را ببینید.
رانش ژن تغییر در فراوانی اللها از نسلی به نسل بعد است؛ که به علت خطای نمونهگیری رخ میدهد.[81] در نتیجه زمانی که نیروهای انتخابکننده غایب یا ضعیف هستند، فراوانی اللها بهطور تصادفی در جهت افزایش یا کاهش «شیفت» پیدا میکند.
شارش ژن
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، شارش ژن، دورگه و انتقال ژن افقی را ببینید.
شارش ژن به معنای تبادل ژنها، خواه بین جمعیتها و خواه بین گونهها است.[82] بود و نبود شارش مسیر تکامل را از اساس تغییر میدهد. پیچیدگی جانداران موجب میشود هر دو جمعیت ایزولهای نهایتاً با سازوکاری خنثی ناسازگاریهای ژنتیکی بیابند. بر اساس مدل بتسون-دوبژانسکی-مولر این ناسازگاری حتی زمانی هم که هر دو جمعیت به گونهای یکسان با محیط سازش یابند؛ رخ میدهد.
سفر رایگان ژنتیکی
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، سفر رایگان ژنتیکی، اثر هیل-رابرتسون و رفت و برگشت انتخابی را ببینید.
نوترکیبی به اللهایی از یک رشته DNA امکان جداشدن میبخشد. البته نرخ نوترکیبی پایین است (تقریباً دو بار به ازای هر کروموزوم در هر نسل). در نتیجه لوکوسهای نزدیک به هم روی یک کروموزوم ممکن است همیشه از هم دور نیفتند؛ و لوکوسهای نزدیک به هم تمایل دارند با هم به ارث برسند پدیدهای که به آن اتصال(به انگلیسی: Linkage) اطلاق شدهاست.[83]
تورش ناشی از جهش
جهش تنها منبعی بزرگ برای گوناگونی ژنی نیست؛ بلکه هرگاه احتمال بروز جهشهای مختلف در سطح مولکولی متفاوت باشد، در تکامل شرکت میکند؛ پدیدهای که به تورش ناشی از جهش شهرت یافتهاست.[84] اگر دو ژنوتیپ برای مثال یکی با نوکلئوتید G و دیگری با نوکلئوتید A، در یک موقعیت یکسان و با ارزش تکاملی یکسان، وجود داشته باشند؛ اما جهشهای G به A بیش از جهشهای A به G رخ دهند؛ آنگاه ژنوتیپ توأم با نوکلئوتید A، تمایل بیشتری به تکامل خواهد داشت.[85]
پیامدها
تکامل هر جنبهای از شکل و رفتار موجودات زنده را تحت تأثیر قرار میدهد. برجستهتر از همه سازگاری رفتاری و جسمی خاصی است که در نتیجه انتخاب طبیعی بهدست میآید. این سازشها با کمک فعالیتهایی از قبیل پیدا کردن غذا، اجتناب از شکارچیان یا جذب جفت؛ تناسب را افزایش میدهند. جانداران همچنین میتوانند با کمک به بستگان خود یا شرکت در همزیستی که برای هر دو طرف سودمند است؛ به انتخاب پاسخ دهند. در دراز مدت، تکامل موجب تقسیم جمعیتهای اجدادی به گونههایی جدید میشود که آمیزش نمیکنند.[86]
این پیامدهای تکامل گاهی به تکامل کلان و تکامل خرد تقسیم میشوند. تکامل کلان در سطح گونهها یا بالاتر از گونهها رخ میدهد، همچون انقراض و گونهزایی؛ در حالی که تکامل خرد تغییرات کوچکتری چون سازش درون یک گونه یا جمعیت را در بر میگیرد.[86] بهطور کلی، تکامل کلان به عنوان نتیجه درازمدت تکامل خرد لحاظ میشود.[87]؛ بنابراین، تمایز تکامل خرد و کلان بنیادین نیست، ناشی از اثر زمان است.[88] اگرچه در تکامل کلان صفات تمام گونهها ممکن است مهم باشد. به عنوان مثال، حجم زیادی از تنوع در میان افراد اجازه میدهد تا یک گونه به سرعت خود را به زیستگاههای جدید وفق دهد، و احتمال انقراض کاهش یابد، در حالی که یک محدوده جغرافیایی گسترده شانس گونهزایی را افزایش میدهد، چون بیشتر احتمال دارد که بخشی از جمعیت جدا و ایزوله شود. در این معنا، تکامل خرد و کلان ممکن است شامل سطوح مختلف انتخاب باشند - تکامل خرد بر ژنها و جانداران عمل کند، در مقابل فرایندهای تکامل کلان مانند انتخاب گونه بر روی کل گونه تأثیر بگذارد و نرخ گونهزایی و انقراض را تغییر بدهد.[89][90][91]
یک تصور غلط رایج این است که تکامل اهداف و برنامههای بلند مدت دارد، واقعیت اینست که تکامل هیچ هدف بلندمدتی ندارد و لزوماً به پیچیدگی بیشتر منجر نمیشود.[92][93] اگر چه گونههای پیچیده تکامل یافتهاند، آنها به عنوان یک اثر جانبی از افزایش تعداد تمام جانداران تلقی میشوند و اشکال ساده حیات همچنان در بیوسفر فراوانتر هستند.[94] اکثریت قریب به اتفاق گونهها پروکاریوتهای میکروسکوپی هستند، که به رغم اندازه کوچکشان نیمی از زیستتوده[95] و اکثریت مطلق تنوع زیستی زمین را تشکیل میدهند.[96] موجودات ساده شکل غالب زندگی روی زمین در طول تاریخ حیات و تا به امروز هستند؛ زندگی پیچیده تنها به این دلیل متنوعتر به نظر میرسد بیشتر قابل توجه است.[97] در واقع، تکامل میکروجانداران به ویژه در پژوهشهای تکاملی مدرن مهم است، چرا که تولید مثل سریع آنها مطالعه تکامل تجربی، و مشاهده تکامل و سازش را در دورههای زمانی کوتاه ممکن میکند.[98][99]
سازش
سازگاری فرایند است که باعث میشود موجودات زنده وفاق بیشتری با زیستگاه خود بیابند.[100][101] همچنین، اصطلاح سازگاری ممکن است به یک صفت که برای بقای جاندار مهم است اشاره کند. به عنوان مثال، سازش دندان اسب برای ساییدن علف. با استفاده از اصطلاح سازش برای اشاره به روند تکامل، و صفت سازشی برای محصول تکامل (بخشی از بدن یا عملکرد بدن) است، این دو معنای کلمه متمایز میشوند. سازشها توسط انتخاب طبیعی تولید میشوند.[102]
همتکاملی
تعامل جانداران میتواند هم ناسازگاری و هم همکاری بیافریند. هر زمان که تعامل میان یک جفت گونه باشد، مثلاً میان پاتوژن و میزبان یا میان شکارچی و شکار، مجموعههای متناظری از سازش توسعه مییابند. بهطوریکه تکامل یک گونه، سازشهایی در گونه دوم موجب میشود. این تغییرات در گونه دوم، در عوض سازشهای جدیدی در گونه اول ایجاد میکند. این چرخه انتخاب و پاسخ همتکاملی نامیده میشود.[103]
همکاری تکاملی
همه فعل و انفعالات تکاملی بین گونهها شامل مناقشه و ستیز نیستند.[104] موارد بسیاری بر اساس منافع متقابل تکامل یافتهاند. برای مثال، میتوان به همکاری شدید بین گیاهان و قارچهای مایکوریزا اشاره کرد.[105]
گونهزایی
گونهزایی فرایندی است که در آن یک گونه به دو یا بیشتر از دو گونه واگرایش مییابد.[106] راههای متعددی برای تعریف مفهوم «گونه» وجود دارد. انتخاب تعریف وابسته به خصوصیات گونه مربوطه است.[107] به عنوان مثال، برخی از مفاهیم گونه به آسانی با توجه به تولیدمثل جنسی جاندار اعمال میشود؛ در حالی که دیگران با عنایت به تولیدمثل غیرجنسی. با وجود اختلاف نظر در مفهوم گونه، این مفاهیم مختلف را میتوان در یکی از سه رویکرد فلسفی زیر گنجاند: بومشناختی، تبارزایشی، یا بر محور جفتگیری.[108]
انقراض
انقراض ناپدید شدن تمام و کمال گونهها است. انقراض یک رویداد غیرمعمول نیست، بهطوریکه گونهها بهطور منظم از طریق گونهزایی ظاهر میشوند و از طریق انقراض ناپدید میشوند.[109] تقریباً تمام گونههای جانوری و گیاهی که زمانی بر روی زمین زندگی میکردهاند اکنون منقرض شدهاند؛ ,[110] و انقراض، به نظر میرسد سرنوشت نهایی همه گونههاست.[111]
تاریخ تکاملی حیات
منشأ حیات
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، آبیوژنز و نظریه جهان RNA را ببینید.
به نظر میرسد که شیمی انرژی بالا ۴ میلیارد سال پیش یک مولکول خود همانندساز تولید کردهاست و آخرین جد مشترک تمام حیات نیم میلیارد سال بعد از آن وجود داشتهاست.[112] اجماع علمی در حال حاضر این است که بیوشیمی پیچیده حیات از برخی سادهترین واکنشهای شیمیایی پدیدار شدهاست.[113] ابتدای حیات احتمالاً شامل مولکول خود همانندسازی چون RNA[114] و مونتاژ سلولهای ساده بودهاست.[115]
نسب مشترک
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، نسب مشترک و شواهد نسب مشترک را ببینید.
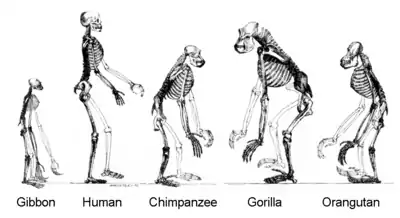
تمام موجودات زنده روی زمین از یک جد مشترک یا یک استخر اجدادی ژن مشتق شدهاند.[116][117] گونههای کنونی تنها یک مرحله در روند تکامل هستند، و تنوع آنها محصول مجموعهای طولانی از گونهزایی و انقراض است.[118]
تکامل حیات
برای جزئیات بیشتر دربارهٔ این بخش از مقاله، تاریخ تکاملی حیات و گاهشمار تکامل را ببینید.
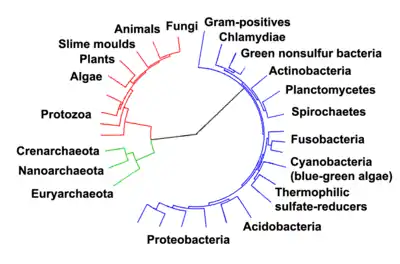
پروکاریوتها از حدود ۳–۴ میلیارد سال پیش ساکنان زمین شدند.[119][120] تغییر واضحی در مورفولوژی یا سازمان سلولی این موجودات طی چند میلیارد سال بعد رخ نداد.[121] سلولهای یوکاریوتی بین ۱٫۶–۲٫۷ میلیارد سال قبل ظاهر شدهاند.
زندگی تا حدود ۶۱۰ میلیون سال پیش شامل یوکاریوتهای تک سلولی، پروکاریوتها و آرکیاها بود تا اینکه در دوره ادیاکاران موجودات چند سلولی در اقیانوسها ظهور یافتند.[119][122]
مدت کمی پس از ظهور اولیه این موجودات چند سلولی، حجم قابل توجهی تنوع زیستی طی حدود ۱۰ میلیون سال، در رویدادی که به نام انفجار کامبرین خوانده میشود پدید آمد.
حدود ۵۰۰ میلیون سال پیش، گیاهان و قارچها در خشکی تکثیر یافتند و به زودی به دنبال آنها بندپایان و جانوران دیگر از راه رسیدند.[123] حشرات به خصوص موفق بودند و حتی امروز عمده گونههای جانوری را تشکیل میدهند.[124] دوزیستان برای اولین بار در حدود ۳۶۴ میلیون سال پیش ظاهر گشتند، به دنبال آنها آمنیوتهای اولیه و پرندگان حدود ۱۵۵ میلیون سال پیش؛ هر دو از تبارهای خزنده مانند، مشتق شدند. حیات پستانداران از حدود ۱۲۹ میلیون سال پیش، نخستیها از ۸۵ میلیون سال پیش، کپیها از ۲۵ میلیون سال پیش، انسانساییان از ۱۳ میلیون سال پیش، سرده انسان از ۲٫۵ میلیون سال پیش و انسان مدرن از حدود ۲۵۰٬۰۰۰ سال پیش آغاز میشود.[125][126][127] با این حال، علیرغم تکامل این جانوران بزرگ، موجودات کوچک شبیه به انواعی که در اوایل این فرایند تکامل یافتند همچنان با موفقیت بسیار به تسلط بر زمین ادامه میدهند؛ چراکه هم بخش اعظم زیستتوده و هم عمده گونهها پروکاریوت هستند.[96]
اعتبار و حمایت علمی
اکثریت گستردهای از جامعه علمی و آکادمیک از تکامل به عنوان تنها توضیح کاملی که میتواند مشاهدات گوناگون در زمینههای زیستشناسی، ژنتیک، زیستشناسی مولکولی، دیرینشناسی، انسانشناسی و… را توضیح دهد، حمایت میکنند.[128][129][130][131][132] تخمینی در سال ۱۹۸۷ بیان کرد که تنها ۷۰۰ نفر از مجموع ۴۸۰۰۰۰ نفر دانشمندان علوم زیستی و زمینشناسی به خلقت گرایی اعتقاد دارند.[133]
پروفسور برایان الترز نویسنده و متخصص در زمینه مناقشه بین خلقتگرایی و تکامل بیان میدارد که ۹۹٫۹ درصد از دانشمندان تکامل را قبول دارند.[134] نظرسنجی که توسط مؤسسه گالوپ در سال ۱۹۹۱ انجام شد نشان داد حدود ۵ درصد از دانشمندان آمریکایی شامل آنهایی که خارج از زمینه علوم زیستی آموزش دیدهاند به خلقتگرایی اعتقاد دارند.[135][136] گرچه در کشوری مانند فرانسه پذیرش نظریه فرگشت تا سال ۱۹۵۰ به تأخیر افتاد ولی محققی مانند فرانک سالووی دلیل این تأخیر را وجود بالای نخستزادگان طی آن دوران در جامعه علمی فرانسه و تمایل آنها به مخالفت با انقلابهای علمی دگراندیشانه نسبت میدهد و نه به ضعف در شواهد تأییدکننده نظریهٔ فرگشت.[137]
به گفتهٔ جیمز واتسون در مقدمهٔ کتابش "ژنتیک مولکولی"، با وجود مخالفتها نظریهٔ فرگشت (تکامل) در دهه ۱۸۶۰ اثبات شد ولی بسیاری همچنان بهشدت به مخالفتهای خود ادامه میدادند که مخالفتهای آنان منشأ علمی نداشته و ناشی از تعصبات و باورهای مذهبی است.[138]
توجیه ژنتیکی فرگشت
امروزه با پیشرفت علم ژنتیک و بیوتکنولوژی، قابلیت مقایسه میان DNA افراد و موجودات از طریق نشانگرهای مولکولی فراهم گشتهاست. از این قابلیت برای شناسایی قرابت و شباهت موجودات با یکدیگر، همچنین برای جرمشناسی و تشخیص والد و فرزند در مسائل قضایی و حقوقی استفاده میشود؛ آزمایش DNA نشان دادهاست که تفاوت میان DNA انسان و شامپانزه در حدود یک درصد است و این خود مهر تأییدی بر نظریهٔ فرگشت (تکامل) میباشد.[139][140][141]
کاربردها
مفاهیم و مدلهای تکامل همچون انتخاب طبیعی کاربردهای بسیاری دارند.[142]
انتخاب مصنوعی انتخاب عامدانه صفات در جمعیتی از جاندارانست. این همان روشی است که از هزاران سال پیش برای اهلی کردن جانوران و گیاهان مورد استفاده قرار گرفتهاست.[143] انتخاب مصنوعی امروزه به جزئی حیاتی از مهندسی ژنتیک تبدیل شدهاست؛ برای مثال میتوان از نشانگرهای انتخابی همچون ژنهای مقاومت میکروبی برای تغییر DNA استفاده کرد.[144]
فهم تغییرات جانداران طی تکامل ژنهای مسبب اعمال و ساختار بدن را مشخص میکند؛ ژنهایی که ممکن است در اختلالات ژنتیکی انسان دخیل باشند.[145]
در علوم کامپیوتر شبیهسازی تکامل با بهکارگیری الگوریتمهای تکاملی و زندگی مصنوعی در دهه ۱۹۶۰ میلادی آغاز شد؛ و به شبیهسازی انتخاب مصنوعی توسعه یافت.[146] در نتیجه کارهای اینگو رچنبرگ انتخاب مصنوعی بهطور گسترده به عنوان یک روش بهینهسازی به رسمیت شناخته شدهاست. رچنبرگ استراتژیهای تکاملی را در حل مسائل پیچیده مهندسی به کار گرفت.[147]
جستارهای وابسته
- میزان حمایت از نظریه تکاملی
- لامارک
- لامارکیسم
- داروین
- داروینیسم
- تاریخ فلسفه تکامل
- تکامل انسان
- مقدمهای بر نظریه تکامل
- زیستشناسی تکاملی
- فرگشتگرایی
- آفرینشگرایی
- طراحی هوشمند
منابع
- جینز، ویلیام. «کتاب فرگشت و انتخاب طبیعی واقعگرایانه». تهران: نشر بینالملل، ۱۳۹۷.
- لوییس، برنارد. «کتاب فرگشت انسان». تهران: نشر طوبی، ۱۳۹۸.
- Futuyma, Douglas J. (2005). Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc. ISBN 0-87893-187-2.
- Jain, R.; Rivera, M.C.; Lake, J.A. (1999). "Horizontal gene transfer among genomes: the complexity hypothesis". Proc Natl Acad Sci U S A. ۹۶ (۷): ۳۸۰۱–۶. Bibcode:1999PNAS...96.3801J. doi:10.1073/pnas.96.7.3801. PMC 22375. PMID 10097118.
- Richardson, Aaron O. and Jeffrey D. Palmer (2007). "Horizontal gene transfer in plants" (PDF). Journal of Experimental Botany. ۵۸ (۱): ۱–۹. doi:10.1093/jxb/erl148. PMID 17030541. Retrieved ۲۰۱۱-۰۱-۳۱.
- Margulis, Lynn (1998). The symbiotic planet: a new look at evolution. Weidenfeld & Nicolson, London. ISBN 0-465-07271-2.
- Sapp, J. (1994). Evolution by association: a history of symbiosis. Oxford University Press, UK. ISBN 0-19-508821-2.
- "Effects of Genetic Drift". University of California at Berkeley. Archived from the original on ۲۳ مارس ۲۰۱۲. Retrieved February ۲۰۱۱. Check date values in:
|accessdate=(help) - Futuyma, Douglas (1998). Evolutionary Biology. Sinauer Associates. p. Glossary. ISBN 0-87893-189-9.
- Darwin, Charles (1859). "XIV". On The Origin of Species. p. ۵۰۳. ISBN 0801413192. Retrieved ۲۰۱۱-۰۲-۲۷.
- Schopf, J.W. (1999). Cradle of life: the discovery of Earth's earliest fossils. Princeton. ISBN 0-691-00230-4.
- Woese, C. (1998). "The Universal Ancestor". PNAS. ۹۵ (۱۲): ۶۸۵۴–۶۸۵۹. Bibcode:1998PNAS...95.6854W. doi:10.1073/pnas.95.12.6854. PMC 22660. PMID 9618502.
- Theobald, D.L. (2010). "A formal test of the theory of universal common ancestry". Nature. ۴۶۵ (۷۲۹۵): ۲۱۹–۲۲۲. Bibcode:2010Natur.465..219T. doi:10.1038/nature09014. PMID 20463738.
- Doolittle, W.F. (February, 2000). "Uprooting the tree of life" (PDF). Scientific American. ۲۸۲ (۲): ۹۰–۹۵. doi:10.1038/scientificamerican0200-90. PMID 10710791. Archived from the original (PDF) on ۳۱ ژانویه ۲۰۱۱. Retrieved ۲۳ مارس ۲۰۱۲. Check date values in:
|year=(help) - Levin, Harold L. ۲۰۰۵. The Earth through time. 8th ed, Wiley, N.Y. Chapter 6: Life on Earth: what do fossils reveal?
- Bowler, Peter J. (2003). Evolution:The History of an Idea. University of California Press. ISBN 0-520-23693-9.
- Kutschera U, Niklas K (2004). "The modern theory of biological evolution: an expanded synthesis". Naturwissenschaften. ۹۱ (۶): ۲۵۵–۷۶. Bibcode:2004NW.....91..255K. doi:10.1007/s00114-004-0515-y. PMID 15241603.
- "IAP Statement on the Teaching of Evolution" (PDF). The Interacademy Panel on International Issues. 2006. Archived from the original (PDF) on 22 April 2007. Retrieved 2007-04-25. Joint statement issued by the national science academies of 67 countries, including the United Kingdom's انجمن سلطنتی
- Board of Directors, American Association for the Advancement of Science (2006-02-16). "Statement on the Teaching of Evolution" (PDF). American Association for the Advancement of Science. from the world's largest general scientific society
- "Statements from Scientific and Scholarly Organizations". National Center for Science Education.
- Dobzhansky, Theodosius 1973. Nothing in biology makes sense except in the light of evolution. American Biology Teacher 35, 125-129. http://www.2think.org/dobzhansky.shtml
- http://www.loghatnaameh.com/dehkhodaworddetail-7ed969bd666b4537a008cfa124669eb9-fa.html
- رادیو زمانه | دانش و فناوری | چرا تکامل برابر درستی برای Evolution نیست؟
- آشوری، داریوش (۱۳۹۳). تعریفها و مفهوم فرهنگ (ویراست ۳). نشر آگه. ص. ۱۵۳. شابک ۹۷۸۹۶۴۳۲۹۰۱۲۲.
- داریوش آشوری (۱۳۷۴)، فرهنگ علوم انسانی (انگلیسی - فارسی))، تهران
- برآیش هستی از دیدگاه داروین
- شبکه چهار برنامهٔ چهار سوی علم (۱ فروردین ۱۳۹۹). «چهار سوی علم -- نژادهای مختلف سگها». دریافتشده در ۴ فروردین ۱۳۹۹.
«این همون سازگاری و تغییر ناشی از انتخاب طبیعیه» (زمان: ۱۴:۳۰ تا ۱۴:۵۰)، فقط این یک مورد نیست، در طول برنامه همیشه از عبارت «سازگاری و تغییر» استفاده میکند
- https://ur.wikipedia.org/wiki/نظریۂ_ارتقا
- https://sd.wikipedia.org/wiki/ارتقا
- Margulis, R.; Fester (1991). Symbiosis as a source of evolutionary innovation: Speciation and morphogenesis. The MIT Press. p. ۴۷۰. ISBN 0262132699.
- Gould, S.J. (2002). The Structure of Evolutionary Theory. Cambridge: Belknap Press (Harvard University Press). ISBN 0-674-00613-5.
- Ghiselin, Michael T. (September/October 1994). "Nonsense in schoolbooks: 'The Imaginary Lamarck'". The Textbook Letter. The Textbook League. Retrieved ۲۰۰۸-۰۱-۲۳. Check date values in:
|publication-date=(help) - Magner, Lois N. (2002). A History of the Life Sciences (Third ed.). Marcel Dekker, انتشارات سیآرسی. ISBN 978-0-203-91100-6.
- Jablonka, E.; Lamb, M. J. (2007). "Précis of evolution in four dimensions". Behavioural and Brain Sciences. ۳۰: ۳۵۳–۳۹۲. doi:10.1017/S0140525X07002221.
- Burkhardt, F.; Smith, S., eds. (1991). The correspondence of Charles Darwin. ۷. Cambridge: Cambridge University Press. pp. ۱۸۵۸–۱۸۵۹.
- Sulloway, F. J. (2009). "Why Darwin rejected intelligent design". Journal of Biosciences. ۳۴ (۲): ۱۷۳–۱۸۳. doi:10.1007/s12038-009-0020-8.
- Dawkins, R. (1990). Blind Watchmaker. Penguin Books. p. ۳۶۸. ISBN 0140144811.
- Darwin, F. (1909). نسخه آرشیو شده (PDF). Cambridge University Press. p. ۵۳. Archived from the original (PDF) on 16 May 2012. Retrieved 24 March 2012. Text "The foundations of the origin of species, a sketch written in 1942 by Charles Darwin" ignored (help)
- Sober, E. (2009). "Did Darwin write the origin backwards?". Proceedings of the National Academy of Sciences. ۱۰۶ (S1): ۱۰۰۴۸–۱۰۰۵۵. Bibcode:2009PNAS..10610048S. doi:10.1073/pnas.0901109106.
- Mayr, Ernst (2001) What evolution is. Weidenfeld & Nicolson, London. p۱۶۵
- Bowler, Peter J. (2003). Evolution: the history of an idea. Berkeley: University of California Press. pp. ۱۴۵–۱۴۶. ISBN 0-520-23693-9. page 147"
- Sokal RR, Crovello TJ (1970). "The biological species concept: A critical evaluation" (PDF). The American Naturalist. ۱۰۴ (۹۳۶): ۱۲۷–۱۵۳. doi:10.1086/282646. JSTOR 2459191. Archived from the original (PDF) on 15 July 2011. Retrieved 23 March 2012.
- Wallace, A (1858). "On the Tendency of Species to form Varieties and on the Perpetuation of Varieties and Species by Natural Means of Selection". Journal of the Proceedings of the Linnean Society of London. Zoology. ۳ (۲): ۵۳–۶۲. doi:10.1098/rsnr.2006.0171. Retrieved ۲۰۰۷-۰۵-۱۳.
- Encyclopædia Britannica Online
- Liu, Y. S.; Zhou, X. M.; Zhi, M. X.; Li, X. J.; Wan, Q. L. (2009). "Darwin's contributions to genetics" (PDF). J Appl Genet. ۵۰ (۳): ۱۷۷–۱۸۴. Archived from the original (PDF) on 30 March 2012. Retrieved 23 March 2012.
- Weiling F (1991). "Historical study: Johann Gregor Mendel ۱۸۲۲–۱۸۸۴". Am. J. Med. Genet. ۴۰ (۱): 1–۲۵, discussion ۲۶. doi:10.1002/ajmg.1320400103. PMID 1887835.
- Wright, S. Evolution and the Genetics of Populations, Volume 1: Genetic and Biometric Foundations. University Of Chicago Press. p. ۴۸۰. ISBN 0226910385.
- Will Provine (1971). The Origins of Theoretical Population Genetics. University of Chicago Press. ISBN 0-226-68464-4.
- Stamhuis, Meijer and Zevenhuizen. /10439561?dopt=Citation Hugo de Vries on heredity, 1889–1903. Statistics, Mendelian laws, pangenes, mutations. , Isis. 1999 Jun;۹۰(۲):۲۳۸–۶۷.
- Quammen, D. (۲۰۰۶). The reluctant Mr. Darwin: An intimate portrait of Charles Darwin and the making of his theory of evolution. New York, NY: W.W. Norton & Company.
- Bowler, Peter J. (1989). The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society. Baltimore: Johns Hopkins University Press. ISBN 978-0-8018-3888-0.
- Watson, J. D.; Crick, F. H. C. "Molecular structure of nucleic acids: A structure for deoxyribose nucleic acid" (PDF). Nature. ۱۷۱ (۴۳۵۶): ۷۳۷–۷۳۸. Bibcode:1953Natur.171..737W. doi:10.1038/171737a0. PMID 13054692.
- Hennig, W.; Lieberman, B. S. (1999). Phylogenetic systematics (New edition (Mar 1 1999) ed.). University of Illinois Press. p. ۲۸۰. ISBN 0252068149.
- Phylogenetics: Theory and practice of phylogenetic systematics (2nd ed.). Wiley-Blackwell. 2011. p. ۳۹۰. doi:10.1002/9781118017883.fmatter.
- Dobzhansky, T. (1973). "Nothing in biology makes sense except in the light of evolution" (PDF). The American Biology Teacher. ۳۵ (۳): ۱۲۵–۱۲۹.
- Cracraft, J.; Bybee, R. W., eds. (2004). Evolutionary science and society: Educating a new generation. Revised Proceedings of the BSCS, AIBS Symposium. Chicago, IL. Archived from the original on 20 July 2011. Retrieved 23 March 2012.
- Avise, J. C.; Ayala, F. J. (2010). "In the Light of Evolution IV. The Human Condition (introduction)" (PDF). Proceedings of the National Academy of Sciences USA. ۱۰۷ (S2): ۸۸۹۷–۸۹۰۱. doi:10.1073/pnas.100321410.
- Pearson H (2006). "Genetics: what is a gene?". Nature. ۴۴۱ (۷۰۹۲): ۳۹۸–۴۰۱. Bibcode:2006Natur.441..398P. doi:10.1038/441398a. PMID 16724031.
- Visscher PM, Hill WG, Wray NR (2008). "Heritability in the genomics era—concepts and misconceptions". Nat. Rev. Genet. ۹ (۴): ۲۵۵–۶۶. doi:10.1038/nrg2322. PMID 18319743.
- Phillips PC (2008). "Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems". Nat. Rev. Genet. ۹ (۱۱): ۸۵۵–۶۷. doi:10.1038/nrg2452. PMC 2689140. PMID 18852697.
- Wu R, Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits". Nat. Rev. Genet. ۷ (۳): ۲۲۹–۳۷. doi:10.1038/nrg1804. PMID 16485021.
- Bossdorf, O.; Arcuri, D.; Richards, C. L.; Pigliucci, M. (2010). "Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana". Evolutionary Ecology. ۲۴ (۳): ۵۴۱–۵۵۳. doi:10.1007/s10682-010-9372-7. Archived from the original on 6 May 2020. Retrieved 24 March 2012.
- Jablonka, E.; Lamb, M. (2005). Evolution in four dimensions: Genetic, epigenetic, behavioural and symbolic. MIT Press. ISBN 0-262-10107-6.
- Jablonka, E.; Raz, G. (2009). "Transgenerational epigenetic inheritance: Prevalence, mechanisms and implications for the study of heredity and evolution" (PDF). The Quarterly Review of Biology. ۸۴ (۲): ۱۳۱–۱۷۶. doi:10.1086/598822. PMID 19606595.
- Chapman, M. J.; Margulis, L. (1998). "Morphogenesis by symbiogenesis" (PDF). International Microbiology. ۱ (۴): ۳۱۹–۳۲۶. PMID 10943381.
- Wilson, D. S.; Wilson, E. O. (2007). "Rethinking the theoretical foundation of sociobiology" (PDF). The Quarterly Review of Biology. ۸۲ (۴). Archived from the original (PDF) on 11 May 2011. Retrieved 24 March 2012.
- Wu R, Lin M (2006). "Functional mapping – how to map and study the genetic architecture of dynamic complex traits". Nat. Rev. Genet. 7 (3): 229–37. doi:10.1038/nrg1804. PMID 16485021.
- Harwood AJ; Harwood, J (1998). "Factors affecting levels of genetic diversity in natural populations". Philos. Trans. R. Soc. Lond. , B, Biol. Sci. 353 (1366): 177–86. doi:10.1098/rstb.1998.0200. PMC 1692205. PMID 9533122
- Ewens W.J. (2004). Mathematical Population Genetics (2nd Edition). Springer-Verlag, New York. ISBN 0-387-20191-2.
- Sawyer SA, Parsch J, Zhang Z, Hartl DL (2007). "Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila". Proc. Natl. Acad. Sci. U.S.A. 104 (16): 6504–10. Bibcode 2007PNAS..104.6504S. doi:10.1073/pnas.0701572104. PMC 1871816. PMID 17409186.
- Hastings, P J; Lupski, JR; Rosenberg, SM; Ira, G (2009). "Mechanisms of change in gene copy number". Nature Reviews. Genetics 10 (8): 551–564. doi:10.1038/nrg2593. PMC 2864001. PMID 1959753
- Carroll SB, Grenier J, Weatherbee SD (2005). From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. Second Edition. Oxford: Blackwell Publishing. ISBN 1-4051-1950-0
- Bowmaker JK (1998). "Evolution of colour vision in vertebrates". Eye (London, England) 12 (Pt 3b): 541–7. doi:10.1038/eye.1998.143. PMID 9775215.
- Radding C (1982). "Homologous pairing and strand exchange in genetic recombination". Annu. Rev. Genet. 16 (1): 405–37. doi:10.1146/annurev.ge.16.120182.002201. PMID 6297377.
- Peters AD, Otto SP (2003). "Liberating genetic variance through sex". BioEssays 25 (6): 533–7. doi:10.1002/bies.10291. PMID 12766942.
- Morjan C, Rieseberg L (2004). "How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles". Mol. Ecol. 13 (6): 1341–56. doi:10.1111/j.1365-294X.2004.02164.x. PMC 2600545. PMID 15140081.
- Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF (2003). "Lateral gene transfer and the origins of prokaryotic groups". Annu Rev Genet 37 (1): 283–328. doi:10.1146/annurev.genet.37.050503.084247. PMID 14616063.
- Baldo A, McClure M (September 1, 1999). "Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts". J. Virol. 73 (9): 7710–21. PMC 104298. PMID 10438861.
- River, M. C. and Lake, J. A. (2004). "The ring of life provides evidence for a genome fusion origin of eukaryotes". Nature 431 (9): 152–5. Bibcode 2004Natur.431..152R. doi:10.1038/nature02848. PMID 15356622.
- Hurst LD (2009). "Fundamental concepts in genetics: genetics and the understanding of selection". Nature Reviews Genetics. 10 (2): 83–93. doi:10.1038/nrg2506. PMID 19119264.
- Orr, H. Allen (August 2009). "Fitness and its role in evolutionary genetics". Nature Reviews Genetics. London: Nature Publishing Group. 10 (8): 531–539. doi:10.1038/nrg2603. ISSN 1471-0056. PMC 2753274. PMID 19546856.
- Haldane, J. B. S. (March 14, 1959). "The Theory of Natural Selection To-Day". Nature. London: Nature Publishing Group. 183 (4663): 710–713. Bibcode:1959Natur.183..710H. doi:10.1038/183710a0. ISSN 0028-0836. PMID 13644170.
- Masel J (2011). "Genetic drift". Current Biology. 21 (20): R837–R838. doi:10.1016/j.cub.2011.08.007. PMID 22032182.
- Morjan, Carrie L.; Rieseberg, Loren H. (June 2004). "How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles". Molecular Ecology. Hoboken, NJ: Wiley-Blackwell. 13 (6): 1341–1356. doi:10.1111/j.1365-294X.2004.02164.x. ISSN 0962-1083. PMC 2600545. PMID 15140081.
- Lien S, Szyda J, Schechinger B, Rappold G, Arnheim N (2000). "Evidence for heterogeneity in recombination in the human pseudoautosomal region: high resolution analysis by sperm typing and radiation-hybrid mapping". Am. J. Hum. Genet. 66 (2): 557–66. doi:10.1086/302754. PMC 1288109. PMID 10677316.
- Lynch, M. (2007). "The frailty of adaptive hypotheses for the origins of organismal complexity". PNAS. 104 (suppl. 1): 8597–8604. Bibcode:2007PNAS..104.8597L. doi:10.1073/pnas.0702207104. PMC 1876435. PMID 17494740.
- Smith N.G.C. , Webster M.T. , Ellegren, H. (2002). "Deterministic Mutation Rate Variation in the Human Genome". Genome Research. 12 (9): 1350–1356. doi:10.1101/gr.220502. PMC 186654. PMID 12213772.
- Scott EC, Matzke NJ (2007). "Biological design in science classrooms". Proc. Natl. Acad. Sci. U.S.A. 104 (suppl_1): 8669–76. Bibcode:2007PNAS..104.8669S. doi:10.1073/pnas.0701505104. PMC 1876445. PMID 17494747.
- Hendry AP, Kinnison MT (2001). "An introduction to microevolution: rate, pattern, process". Genetica. 112–113: 1–8. doi:10.1023/A:1013368628607. PMID 11838760.
- Leroi AM (2000). "The scale independence of evolution". Evol. Dev. 2 (2): 67–77. doi:10.1046/j.1525-142x.2000.00044.x. PMID 11258392.
- Gould 2002, pp. 657–658.
- Gould SJ (1994). "Tempo and mode in the macroevolutionary reconstruction of Darwinism". Proc. Natl. Acad. Sci. U.S.A. 91 (15): 6764–71. Bibcode:1994PNAS...91.6764G. doi:10.1073/pnas.91.15.6764. PMC 44281. PMID 8041695.
- Jablonski, D. (2000). "Micro- and macroevolution: scale and hierarchy in evolutionary biology and paleobiology". Paleobiology. 26 (sp4): 15–52. doi:10.1666/0094-8373(2000)26[15:MAMSAH]2.0.CO;2.
- Michael J. Dougherty. Is the human race evolving or devolving? ساینتیفیک آمریکن July 20, 1998.
- TalkOrigins Archive response to آفرینشگرایی claims – Claim CB932: Evolution of degenerate forms
- Carroll SB (2001). "Chance and necessity: the evolution of morphological complexity and diversity". Nature. 409 (6823): 1102–9. doi:10.1038/35059227. PMID 11234024.
- Whitman W, Coleman D, Wiebe W (1998). "Prokaryotes: the unseen majority". Proc Natl Acad Sci U S A. 95 (12): 6578–83. Bibcode:1998PNAS...95.6578W. doi:10.1073/pnas.95.12.6578. PMC 33863. PMID 9618454.
- Schloss P, Handelsman J (2004). "Status of the microbial census". Microbiol Mol Biol Rev. 68 (4): 686–91. doi:10.1128/MMBR.68.4.686-691.2004. PMC 539005. PMID 15590780.
- Nealson K (1999). "Post-Viking microbiology: new approaches, new data, new insights". Orig Life Evol Biosph. 29 (1): 73–93. doi:10.1023/A:1006515817767. PMID 11536899.
- Buckling A, Craig Maclean R, Brockhurst MA, Colegrave N (2009). "The Beagle in a bottle". Nature. 457 (7231): 824–9. Bibcode:2009Natur.457..824B. doi:10.1038/nature07892. PMID 19212400.
- Elena SF, Lenski RE (2003). "Evolution experiments with microorganisms: the dynamics and genetic bases of adaptation". Nature Reviews Genetics. 4 (6): 457–69. doi:10.1038/nrg1088. PMID 12776215.
- Mayr, Ernst 1982. The growth of biological thought. Harvard. p483: "Adaptation... could no longer be considered a static condition, a product of a creative past and became instead a continuing dynamic process."
- The Oxford Dictionary of Science defines adaptation as "Any change in the structure or functioning of an organism that makes it better suited to its environment".
- Orr H (2005). "The genetic theory of adaptation: a brief history". Nature Reviews Genetics. 6 (2): 119–27. doi:10.1038/nrg1523. PMID 15716908.
- Wade MJ (2007). "The co-evolutionary genetics of ecological communities". Nature Reviews Genetics. 8 (3): 185–95. doi:10.1038/nrg2031. PMID 17279094.
- Sachs J (2006). "Cooperation within and among species". J. Evol. Biol. 19 (5): 1415–8, discussion 1426–36. doi:10.1111/j.1420-9101.2006.01152.x. PMID 16910971.
*Nowak M (2006). "Five rules for the evolution of cooperation". Science. 314 (5805): 1560–3. Bibcode:2006Sci...314.1560N. doi:10.1126/science.1133755. PMC 3279745. PMID 17158317. - Paszkowski U (2006). "Mutualism and parasitism: the yin and yang of plant symbioses". Curr. Opin. Plant Biol. 9 (4): 364–70. doi:10.1016/j.pbi.2006.05.008. PMID 16713732.
- Gavrilets S (2003). "Perspective: models of speciation: what have we learned in 40 years?". Evolution. 57 (10): 2197–215. doi:10.1554/02-727. PMID 14628909.
- de Queiroz K (2005). "Ernst Mayr and the modern concept of species". Proc. Natl. Acad. Sci. U.S.A. 102 (Suppl 1): 6600–7. Bibcode:2005PNAS..102.6600D. doi:10.1073/pnas.0502030102. PMC 1131873. PMID 15851674.
- Ereshefsky, M. (1992). "Eliminative pluralism". Philosophy of Science. 59 (4): 671–690. doi:10.1086/289701. JSTOR 188136.
- Benton MJ (1995). "Diversification and extinction in the history of life". Science. 268 (5207): 52–8. Bibcode:1995Sci...268...52B. doi:10.1126/science.7701342. PMID 7701342.
- Raup DM (1986). "Biological extinction in Earth history". Science. 231 (4745): 1528–33. Bibcode:1986Sci...231.1528R. doi:10.1126/science.11542058. PMID 11542058.
- Avise JC, Hubbell SP, Ayala FJ. (2008). "In the light of evolution II: Biodiversity and extinction". Proc. Natl. Acad. Sci. U.S.A. 105 (Suppl 1): 11453–7. Bibcode:2008PNAS..10511453A. doi:10.1073/pnas.0802504105. PMC 2556414. PMID 18695213.
- Doolittle, W. Ford (2000). "Uprooting the tree of life". Scientific American. 282 (6): 90–95. doi:10.1038/scientificamerican0200-90. PMID 10710791. Unknown parameter
|month=ignored (help) - Peretó J (2005). "Controversies on the origin of life" (PDF). Int. Microbiol. 8 (1): 23–31. PMID 15906258.
- Joyce GF (2002). "The antiquity of RNA-based evolution". Nature. 418 (6894): 214–21. Bibcode:2002Natur.418..214J. doi:10.1038/418214a. PMID 12110897.
- Trevors JT, Psenner R (2001). "From self-assembly of life to present-day bacteria: a possible role for nanocells". FEMS Microbiol. Rev. 25 (5): 573–82. doi:10.1111/j.1574-6976.2001.tb00592.x. PMID 11742692.
- Penny D, Poole A (1999). "The nature of the last universal common ancestor". Curr. Opin. Genet. Dev. 9 (6): 672–77. doi:10.1016/S0959-437X(99)00020-9. PMID 10607605.
- Theobald, DL. (2010). "A formal test of the theory of universal common ancestry". Nature. 465 (7295): 219–22. Bibcode:2010Natur.465..219T. doi:10.1038/nature09014. PMID 20463738.
- Bapteste E, Walsh DA (2005). "Does the 'Ring of Life' ring true?". Trends Microbiol. 13 (6): 256–61. doi:10.1016/j.tim.2005.03.012. PMID 15936656.
- Cavalier-Smith T (2006). "Cell evolution and Earth history: stasis and revolution". Philosophical Transactions of the Royal Society B. 361 (1470): 969–1006. doi:10.1098/rstb.2006.1842. PMC 1578732. PMID 16754610.
- Schopf J (2006). "Fossil evidence of Archaean life". Philosophical Transactions of the Royal Society B. 361 (1470): 869–85. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
*Altermann W, Kazmierczak J (2003). "Archean microfossils: a reappraisal of early life on Earth". Res Microbiol. 154 (9): 611–17. doi:10.1016/j.resmic.2003.08.006. PMID 14596897. - Schopf J (1994). "Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic". Proc Natl Acad Sci U S A. 91 (15): 6735–42. Bibcode:1994PNAS...91.6735S. doi:10.1073/pnas.91.15.6735. PMC 44277. PMID 8041691.
- DeLong E, Pace N (2001). "Environmental diversity of bacteria and archaea". Syst Biol. 50 (4): 470–8. doi:10.1080/106351501750435040. PMID 12116647.
- Waters ER (2003). "Molecular adaptation and the origin of land plants". Mol. Phylogenet. Evol. 29 (3): 456–63. doi:10.1016/j.ympev.2003.07.018. PMID 14615186.
- Mayhew PJ (2007). "Why are there so many insect species? Perspectives from fossils and phylogenies". Biol Rev Camb Philos Soc. 82 (3): 425–54. doi:10.1111/j.1469-185X.2007.00018.x. PMID 17624962.
- Carroll, Robert L. (May 2007). "The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians". Zoological Journal of the Linnean Society. 150 (Supplement s1): 1–140. doi:10.1111/j.1096-3642.2007.00246.x. ISSN 1096-3642.
- Wible JR, Rougier GW, Novacek MJ, Asher RJ (2007). "Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary". Nature. 447 (7147): 1003–1006. Bibcode:2007Natur.447.1003W. doi:10.1038/nature05854. PMID 17581585.
- Witmer LM (2011). "Palaeontology: An icon knocked from its perch". Nature. 475 (7357): 458–459. doi:10.1038/475458a. PMID 21796198.
- Myers, PZ (2006-06-18). "Ann Coulter: No evidence for evolution?". Pharyngula. scienceblogs.com. Archived from the original on 22 June 2006. Retrieved 2006-11-18.
- The National Science Teachers Association's position statement on the teaching of evolution. بایگانیشده در ۱۹ آوریل ۲۰۰۳ توسط Wayback Machine
- IAP Statement on the Teaching of Evolution بایگانیشده در ۱۷ ژوئیه ۲۰۱۱ توسط Wayback Machine Joint statement issued by the national science academies of 67 countries, including the United Kingdom's انجمن سلطنتی (PDF file)
- From the انجمن پیشبرد علوم آمریکا، the world's largest general scientific society: 2006 Statement on the Teaching of Evolution (PDF file), AAAS Denounces Anti-Evolution Laws
- <520%3AFFAMOH>2.0.CO3B2-P Fact, Fancy, and Myth on Human Evolution, Alan J. Almquist, John E. Cronin, Current Anthropology, Vol. 29, No. 3 (Jun. , 1988), pp. 520–522
- As reported by Newsweek: "By one count there are some 700 scientists with respectable academic credentials (out of a total of 480,000 U.S. earth and life scientists) who give credence to creation-science, the general theory that complex life forms did not evolve but appeared 'abruptly'."Martz & McDaniel 1987, p. 23
- Finding the Evolution in Medicine بایگانیشده در ۲۲ نوامبر ۲۰۰۸ توسط Wayback Machine, Cynthia Delgado, NIH Record, July 28, 2006.
- Public beliefs about evolution and creation, Robinson, B. A. 1995.
- Many scientists see God's hand in evolution, Witham, Larry, Reports of the National Center for Science Education 17(6): 33, 1997
- سالووی، فرانک (۱۳۸۶). دگراندیشان شورشی. ۱. ترجمهٔ حسینعلی مقیمی. تهران: نشر دایره. ص. ۷۹. شابک ۹۷۸۹۶۴۶۸۳۹۷۳۱.
- واتسون، جیمز (۲۰۰۸). ژنتیک مولکولی. تهران: خانهی زیستشناسی. صص. مقدمه. شابک ۹۷۸-۹۶۴-۲۶۰۵-۶۲-۰.
- "Genomewide Comparison of DNA Sequences between Humans and Chimpanzees". The American Journal of Human Genetics. 70 (6): 1490–1497. 2002-06-01. doi:10.1086/340787. ISSN 0002-9297.
- Wong, Kate (2014-08-19). "The 1 Percent Difference". Scientific American. 311 (3): 100–100. doi:10.1038/scientificamerican0914-100. ISSN 0036-8733.
- «2005 Release: New Genome Comparison Finds Chimps, Humans Very Similar at DNA Level». National Human Genome Research Institute (NHGRI) (به انگلیسی). دریافتشده در ۲۰۱۸-۰۸-۲۴.
- Bull JJ, Wichman HA (2001). "Applied evolution". Annu Rev Ecol Syst. 32 (1): 183–217. doi:10.1146/annurev.ecolsys.32.081501.114020.
- Doebley JF, Gaut BS, Smith BD (2006). "The molecular genetics of crop domestication". Cell. 127 (7): 1309–21. doi:10.1016/j.cell.2006.12.006. PMID 17190597.
- Jäckel C, Kast P, Hilvert D (2008). "Protein design by directed evolution". Annu Rev Biophys. 37 (1): 153–73. doi:10.1146/annurev.biophys.37.032807.125832. PMID 18573077.
- Maher B. (2009). "Evolution: Biology's next top model?". Nature. 458 (7239): 695–8. doi:10.1038/458695a. PMID 19360058.
- Fraser AS (1958). "Monte Carlo analyses of genetic models". Nature. 181 (4603): 208–9. Bibcode:1958Natur.181..208F. doi:10.1038/181208a0. PMID 13504138.
- Rechenberg, Ingo (1973). Evolutionsstrategie – Optimierung technischer Systeme nach Prinzipien der biologischen Evolution (PhD thesis) (به German). Fromman-Holzboog.
پیوند به بیرون
| در ویکیانبار پروندههایی دربارهٔ فرگشت موجود است. |
- ویدیوهای علمی فرگشت در وبسایت «تد دات کام» (همراه با زیرنویس فارسی)
- سلسله مقالات علمی ترجمه شده درباره فرگشت، در وبگاه رادیو زمانه
- Understanding Evolution - a guide prepared by the University of California at Berkeley
- Howstuffworks.com—How Evolution Works
- The Big Picture on Evolution by the Welcome Trust.
- Online collection of Darwin's papers at the University of Cambridge
- Talk Origins in depth website on information about evoluton and the evidence for it
- National Center for Science Education Information on how evolution works
- PBS, on evolution site
