ژن
ژن (به فرانسوی: gène) در ژنتیک مندلی و ژنتیک مولکولی کلاسیک، همچنین در مبحث تکامل و در دوران پساژنگان تعاریف متفاوتی دارد. اما از دیدگاه مولکولی کلاسیک دنبالهای از نوکلوئوتیدها است که در برگیرنده اطلاعات لازم جهت تولید مولکولهای RNA یا پروتئینهای لازم برای سلول هستند. هر ژن در بخشی از DNA سلول وجود دارد. یا با تعریفی دیگر ژن بخشی از مولکول دی ان ای میباشد که روی یک رشته از آن قرار دارد.
درون سلولها، طی فرایند رونویسی ژنها به مولکولهای RNA تبدیل میشوند که یا به شکل مستقیم در سلول استفاده میشوند یا دربرگیرنده اطلاعاتی جهت تولید پروتئین هستند و طی فرایند ترجمه، پروتئین مربوط به آنها ساخته میشود.
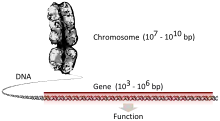
ژنها تمامی صفات سلول را کنترل میکنند و عملکرد سلولها به کمک ژنها و پروتئینهای ساخته شده از روی آنها تعیین میشود. این ژنها از پدر و مادر به ارث میرسند و ممکن است به مرور در اثر فرایند تقسیم سلولی یا در تعامل سلول با محیط بیرونی دچار تغییر شوند.
ژنها میتوانند دچار جهش شوند، که تغییری در توالی دنباله آنهاست. جهشها باعث ایجاد تغییر در پروتئینهای تولید شده از روی ژنها میشوند و میتوانند عملکرد پروتئین را به کلی تغییر دهند و باعث به وجود آمدن صفاتی جدید در سلول بشوند. ژنها در اثر جهشهای ژنتیکی، براساس انتخاب طبیعی تکامل پیدا میکنند و ژنهای قویتر و بهتر به مرور جایگزین ژنهای قبلی میشوند.
واژه ژن نخستین بار در سال ۱۹۰۹، توسط یک گیاهشناس و نسلشناس دانمارکی به نام ویلهلم یوهانسون مطرح شد.[1]
ساختار و عملکرد ژن
ساختار
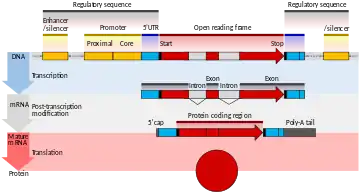
ژن بخشی از یک مولکول نوکلئیک اسید، DNA یا RNA است که یک محصول عملکردی یا فاشینال از روی آن ساخته میشود.
ساختار ژن در برگیرنده تعداد زیادی جز است که رشته تولیدکننده پروتئین بخشی از آن است. این بخشها شامل مناطقی هستند که طی فرایند رونویسی ساخته نمیشوند یا ترجمه روی آنها انجام نمیشود.
هر ژن دارای بخش دنباله نظارتی است که برای بیان ژن ضروری است. این بخش شامل پروموتر است که به وسیله فاکتورهای رونویسی تشخیص داده میشود و به آنها متصل میشود.
این فاکتورها آنزیم تولید RNA را به کار میگیرند و فرایند تولید RNA از روی رشته DNA آغاز میشود. ژنهایی که به مقدار زیاد در سلول استفاده میشوند و به عبارتی بیان بالایی دارند پروموتر قویای دارند و ژنهایی که به ندرت در سلول به کار گرفته میشوند پروموتر ضعیفی دارند.[2]:7.2
بخش افزاینده(Enhancer) نیز موجود است که پروتئینهای فعالکننده به آن میچسبد و میزان بیان ژن را افزایش میدهد. در طرف مقابل بخش خاموشکننده(Silencer) وجود دارد که پروتئینهای خاصی به آن متصل میشوند و از میزان بیان و رونویسی ژن میکاهند.[2]:7.1بدین ترتیب و به وسیله این بخشها، میزان بیان ژنها کنترل میشود. در نهایت آن بخشی از ژن که رونویسی میشود رشته RNA پیام رسان اولیه را تولید میکند. این رشته طی یک فرایند، اینترونها و بخشهایی از ابتدا و انتهایش حذف میشود و با ادغام بخشهای باقیمانده (اگزونها)، رشته بالغ به دست میآید. به این رشته ثانویه، رشته RNA پیام رسان بالغ میگویند.[3]
از روی این رشته ثانویه پروتئینها ساخته میشوند.
عملکرد
تعیین دقیق این موضوع که یک ژن چه عملیاتی را در سلول انجام میدهد، بسیار دشوار است. بخشهای نظارتی یک ژن مثل افزاینده الزاماً در نزدیکی یک ژن نیستند. بعلاوه در خود ژن نیز ممکن است بخشهای اینترون خیلی بزرگ باشند که عملاً یافتن بخش ترجمه شونده ژن را دشوار میکنند.[4]
مطالعات اولیه این تئوری را در اذهان ایجاد کرد که هر ژن تولیدکننده یک پروتئین است و بر این اساس میتوان عملکرد ژنها را ارزیابی کرد. هر چند این مفهوم با کشف اینکه یک ژن میتواند با قطعه قطعهشدنهای مختلف اینترونها، RNAهای پیامرسان متفاوت تولید کند دچار بهبود و تعریف مجدد شد.[5][6]
بیان ژنها
در تمامی ارگانیسمهای زنده، دو گام جهت خواندن اطلاعات موجود در DNA و تبدیل آن اطلاعات به پروتئینهای مخصوص وجود دارد. گام اول رونویسی DNA به رشته RNA پیامرسان است[2]:6.1 و گام دوم ترجمه رشته به پروتئین.[2]:6.2 ژنهایی که پروتئین نمیسازند و خود RNA درون سلول استفاده میشود همچنان گام اول را طی میکنند اما گام دوم برای آنها اجرا نمیشود.[7]
فرایند تولید یک مولکول کاربردی زیستی از روی یک ژن، بیان ژن نام دارد و محصول به دست آمده که یک پروتئین یا RNA است، محصول ژن نامیده میشود.
کد ژنتیکی

دنباله نوکلئوتیدی یک ژن، دنباله آمینواسیدی مربوط به آن را معین میکند. هر بخش سه عضوی از نوکلئوتیدها یک رمز ژنتیکی نامیده میشود که بیانگر دقیقاً یک آمینواسید است.[2]:6 به علاوه درون اطلاعات ژنها یک رمز شروع. یک رمز پایان داریم که شروع و پایان فرایند ترجمه را معلوم میکنند. چون رشته RNA، یک رشته حرفی است، پس مجموعاً رمز مختف وجود دارد. اما تنها ۲۰ مورد آمینواسید شناخته شدهاست، این به این معناست که برخی از رمزها، به آمینواسیدهای یکسانی اشاره میکنند.[8]


رونویسی
این فرایند یک رشته RNA پیامرسان تولید میکند که نوکلئوتیدهایش از حروف A, U, G, C تشکیل شدهاست و مکمل وارون رشته DNA ای است که رونویسی از آن انجام شدهاست.[2]:6.1 این رشته یک لایه میانی میان ژن موجود در DNA و پروتئین مرتبط با آن است. فرایند رونویسی به کمک آنزیمی به نام RNA پلیمراز انجام میگیرد. برای شروع فرایند رونویسی، فاکتورهای رونویسی ابتدا منطقه پروموتر را شناسایی میکنند و به آنها میچسبند و نهایتاً با کمک بخشهای افزاینده و سایز آنزیمهای فعالکننده، آنزیم RNA پلیمراز به کار گرفته میشود و فرایند رونویسی شروع میشود.[2]:7
در سلولهای پروکاریوت که فاقد هسته هستند، این عملیات درون سیتوپلاسم صورت میگیرد. در سلولهای یوکاریوت چون ماده وراثتی در هسته قرار دارد، رونویسی نیز درون هسته صورت میگیرد و ابتدا رشته اولیه را تولید میکند. سپس ترکیبی از اینترونها از این رشته کنده شده و بخشهایی از ابتدا و انتهای آن نیز بریده میشود و رشته RNA پیامرسان نهایی تولید میشود. این رشته از هسته سلول خارج میشود. این فرایند حذف اینترونها وابسته به وضعیت و نیاز سلول میتواند به شکلهای مختلفی انجام شود لذا یک ژن در واقع میتواند تعداد زیادی RNA پیامرسان بالغ متفاوت تولید کند. این موضوع در سلولهای یوکاریوت و برخی جانداران پروکاریوت دیده میشود.[2]:7.5[9]
ترجمه
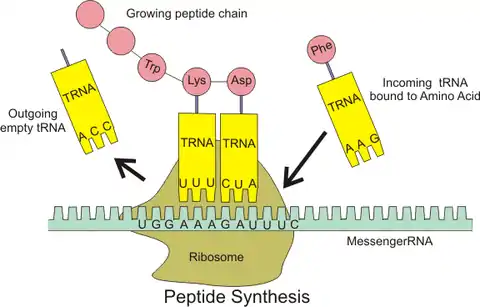
این فرایند از روی رشته RNA پیامرسان بالغ، پروتئین تولید میکند. فرایند ترجمه به کمک پروتئینی بزرگ و پیچیده به نام ریبوزوم انجام میشود. طی این فرایند رشته RNA وارد ریبوزوم میشود و ریبوزوم رمزهای رشته را سه حرف به سه حرف میخواند و با اضافه کردن آمینواسید مربوطه یک دنباله از آمینواسیدها میسازد که پروتئین را تشکیل میدهد. این آمینواسیدها به وسیله پیوند پپتیدی به یکدیگر متصل میشوند. مرحله افزودن آمینواسیدها به کمک رشتههای RNA حامل صورت میگیرد. این RNAها، از یک طرف به آمینواسید متصلند و از طرف دیگر دربرگیرنده مکمل رشته رمز هستند که در ریبوزوم با رشته رمز پیوند میخورند و جذب ریبوزوم میشوند و آمینواسید متصل به آنها توسط ریبوزوم جدا شده و به دنباله آمینواسیدی تولید شده میچسبد.[2]:3
ساماندهی بیان ژنها
ژنها باید ساماندهی شوند، به گونهای که تنها هنگامی که سلول به آنها نیاز دارد بیان شوند. یک سلول میزان بیان یک ژن را بر اساس شرایط محیطی (مانند دما، مواد اولیه موجود و …) و شرایط داخلی (مانند متابولیسم، چرخه سلولی و …) و از همه مهمتر کاربردش در یک ارگانیسم پیچیده تعیین میکند. به عنوان مثال تمامی سلولهای بدن ما DNA و به تبع آن ژنهای یکسانی دارند، اما تفاوت در میزان بیان ژنها به جهت عملکرد متفاوت باعث میشود که سلولهای چشم ما سلولهایی حساس به نور باشند و در طرف مقابل سلولهای روی پوست، سلولهایی مقاوم و با عملکرد متفاوت باشند.[2]:7
ساماندهی در مراحل مختلف تولید پروتئین میتواند انجام گیرد. در گام شروع رونویسی و به کمک فاکتورهای فعالکننده و خاموشکننده، در گام تولید RNA بالغ به کمک روشهای مختلف حذف برخی اینترونها و در گام پس از ترجمه و تغییر در ساختار پروتئین.[10]
تکامل مولکولی
فرایند تقسیم سلولی که با کپی شدن DNA همراه است به جهت وجود ساز و کارهایی جهت بررسی درستی فرایند، عملیاتی بسیار دقیق است،[2]:7.6 طوری که در سلولهای یوکاریوت به ازای یک بار کپیکردن DNA، احتمال خطا در هر نوکلئوتید در حدود است.[11] این خطا میتواند تغییر در یک نقطه از DNA یا اضافه شدن و کم شدن نوکلئوتید در DNA باشد. هر یک از این جهشها میتواند باعث تغییرات در ژنها بشوند. به طوری که یک ژن دیگر کارایی قبلی را نداشته باشد و عملکردش دچار تغییر شود. چون دنباله آمینواسیدی که تولید میکند متفاوت میشود و به تبع آن ساختار پروتئین دچار تفاوت میشود.

اکثر تغییرات در DNA، خنثی هستند و اثری در سلول ندارند که به آنها جهش خاموش میگویند. این جهشها ممکن است باعث تغییر در رمزهای ژنها بشوند اما تغییری در آمینواسیدی که آن ژن تولید میکند نداشته باشند. یا اگر هم باعث تغییر در آمینواسید بشوند، تغییر قابل توجهی در ساختار پروتئین نکنند و پروتئین همچنان کارایی قبلیش را داشته باشد.
برخی از جهشها میتوانند باعث تغییرات زیادی شوند. در اینصورت سلول رفتار متفاوتی نشان میدهد که میتواند مضر باشد و این سلول به کمک انتخاب طبیعی حذف میشود. بخش بسیار کوچکی از جهشها باعث میشوند که ژنهای تغییر یافته مفید باشند و این سلول با یک ژن بهتر همچنان حفظ شود و تولید مثل کند و ژن جدید جایگزین ژن قبلی بشود. به این شکل ژنها دچار تکامل میشوند.[2]:7.6
ژنوم
تمامی ماده وراثتی در یک سلول به عنوان ژنوم شناخته میشود که دربرگیرنده ژنها و دیگر بخشهای DNA است که کاربردی در ساختار پروتئینها ندارند.[12]
تعداد ژنها
سایز ژنوم و تعداد ژنهایی که در خود ذخیره کردهاست در میان جانداران مختلف بسیار متفاوت است. سادهترین و کوچکترین ژنومها مربوط به ویروسهاست که ماده وراثتیشان به فرم یک RNA است.[13] در طرف مقابل گیاهان وجود دارند که گاهی تعداد بسیار کثیری ژن در آنها وجود دارد.[14] تعداد پروتئینهایی که از روی ژنها تولید میشوند در حدود ۵ میلیون ساختار مختلف تخمین زده میشود.[15]
در مورد انسان با گذشت زمان و بیان تعریفی دقیقتر و جامعتر از ژنها، تعداد ژنهای شناخته شده در بدن انسان به تدریج کاهش پیدا کرد و اکنون وجود حدود ۲۰۰۰۰ ژن در DNA انسان تخمین زده میشود.[16] در انسان تنها حدود ۱ الی ۲ درصد کل ژنوم را ژنها تشکیل میدهند.[17] تمامی سلولهای بدن یک جاندار ژنومی کاملاً تشابه دارند اما سلولهای متفاوت از ژنهای مختلفی استفاده میکنند.
منابع
- Johannsen W (1905). Arvelighedslærens elementer [The Elements of Heredity] (به Danish). Copenhagen. Rewritten, enlarged and translated into German as Johannsen W (1909). Elemente der exakten Erblichkeitslehre. Jena: Gustav Fischer. Archived from the original on 30 May 2009. Retrieved 15 May 2020.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2002). Molecular Biology of the Cell (Fourth ed.). New York: Garland Science. ISBN 978-0-8153-3218-3.
- Bicknell AA, Cenik C, Chua HN, Roth FP, Moore MJ (December 2012). "Introns in UTRs: why we should stop ignoring them". BioEssays. 34 (12): 1025–34. doi:10.1002/bies.201200073. PMID 23108796.
- Gericke NM, Hagberg M (5 December 2006). "Definition of historical models of gene function and their relation to students' understanding of genetics". Science & Education. 16 (7–8): 849–881. Bibcode:2007Sc&Ed..16..849G. doi:10.1007/s11191-006-9064-4.
- Marande W, Burger G (October 2007). "Mitochondrial DNA as a genomic jigsaw puzzle". Science. AAAS. 318 (5849): 415. Bibcode:2007Sci...318..415M. doi:10.1126/science.1148033. PMID 17947575.
- Parra G, Reymond A, Dabbouseh N, Dermitzakis ET, Castelo R, Thomson TM, et al. (January 2006). "Tandem chimerism as a means to increase protein complexity in the human genome". Genome Research. 16 (1): 37–44. doi:10.1101/gr.4145906. PMC 1356127. PMID 16344564.
- Eddy SR (December 2001). "Non-coding RNA genes and the modern RNA world". Nature Reviews. Genetics. 2 (12): 919–29. doi:10.1038/35103511. PMID 11733745.
- Crick FH (October 1962). "The genetic code". Scientific American. WH Freeman and Company. 207 (4): 66–74. doi:10.1038/scientificamerican1062-66. PMID 13882204.
- Woodson SA (May 1998). "Ironing out the kinks: splicing and translation in bacteria". Genes & Development. 12 (9): 1243–7. doi:10.1101/gad.12.9.1243. PMID 9573040.
- Jacob F, Monod J (June 1961). "Genetic regulatory mechanisms in the synthesis of proteins". Journal of Molecular Biology. 3 (3): 318–56. doi:10.1016/S0022-2836(61)80072-7. PMID 13718526.
- Nachman MW, Crowell SL (September 2000). "Estimate of the mutation rate per nucleotide in humans". Genetics. 156 (1): 297–304. PMC 1461236. PMID 10978293.
- Ridley, M. (2006). Genome. New York, NY: Harper Perennial. شابک ۰−۰۶−۰۱۹۴۹۷−۹
- Belyi VA, Levine AJ, Skalka AM (December 2010). "Sequences from ancestral single-stranded DNA viruses in vertebrate genomes: the parvoviridae and circoviridae are more than 40 to 50 million years old". Journal of Virology. 84 (23): 12458–62. doi:10.1128/JVI.01789-10. PMC 2976387. PMID 20861255.
- Zonneveld, B.J.M. (2010). "New Record Holders for Maximum Genome Size in Eudicots and Monocots". Journal of Botany. 2010: 1–4. doi:10.1155/2010/527357.
- Perez-Iratxeta C, Palidwor G, Andrade-Navarro MA (December 2007). "Towards completion of the Earth's proteome". EMBO Reports. 8 (12): 1135–41. doi:10.1038/sj.embor.7401117. PMC 2267224. PMID 18059312.
- Pertea M, Salzberg SL (2010). "Between a chicken and a grape: estimating the number of human genes". Genome Biology. 11 (5): 206. doi:10.1186/gb-2010-11-5-206. PMC 2898077. PMID 20441615.
- Claverie JM (September 2005). "Fewer genes, more noncoding RNA". Science. 309 (5740): 1529–30. Bibcode:2005Sci...309.1529C. doi:10.1126/science.1116800. PMID 16141064.
