دستگاه غشایی درونی
دستگاه غشایی درونی از غشاهای گوناگون شناور در سیتوپلاسم یاخته یوکاریوتی تشکیل شدهاست. این غشاها یاخته را به بخشهای کارکردی و ساختاری یا اندامکها تقسیم میکنند. در یوکاریوتها اندامکهای دستگاه غشایی درونی عبارتند از: پوشش هستهای، شبکهٔ آندوپلاسمی، دستگاه گلژی، لیزوزوم، وزیکول، اندوزوم و غشای پلاسما (یاختهای). این دستگاه در یک تعریف دقیقتر، مجموعهای از غشاهاست که یک واحد کارکردی و تکاملی را تشکیل میدهند که مستقیماً به هم پیوند دارند یا مواد را از طریق وزیکولها جابهجا میکنند. نکته مهم این است که غشاهای کلروپلاست یا میتوکندری جزو دستگاه غشایی درونی نیستند؛ گرچه برخی از اجزای این دستگاه از میتوکندری تکامل یافتهاند.
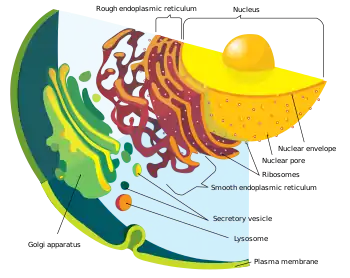
پوشش هستهای از دو لایه غشای لیپیدی دولایهٔ ساخته شدهاست و محتوای هسته را در بر میگیرد. شبکهٔ آندوپلاسمی اندامکی سازنده (سنتزکننده) و انتقالدهنده است که در یاختههای گیاهی و جانوری در داخل سیتوپلاسم منشعب میشود. دستگاه گلژی مجموعهای از چند کیسه است که در آن مولکولها برای تحویل به سایر اجزای یاخته یا ترشح از یاخته بستهبندی میشوند. واکوئلها که هم در یاختههای گیاهی و هم در جانوران یافت میشوند (اگرچه در یاختههای گیاهی بسیار بزرگتر هستند)، مسئول حفظ شکل و ساختار یاخته و همچنین ذخیره مواد زائد هستند. وزیکول کیسهای غشایی نسبتاً کوچک است که مواد را ذخیره یا حمل میکند. اندامکی معروف به اشپیتزنکاپه نیز در قارچها یافت میشود که با رشد نوک نخینه در ارتباط است.
پیشینه مفهوم
در مخمر، بیشتر لیپیدها در شبکه آندوپلاسمی، ذرات لیپید یا میتوکندری ساخته میشوند و ساخت لیپید در غشای پلاسما یا غشای هستهای کم است یا اصلاً وجود ندارد.[1][2] بیوسنتز اسفنگولیپید در شبکه آندوپلاسمی آغاز و در دستگاه گلژی تکمیل میشود.[3] در پستانداران نیز به استثنای چند مرحله نخست بیوسنتز لیپید اتری که در پراکسیزومها رخ میدهد، وضعیت مشابه است.[4] لیپیدها با انتقال از این مکانهای سازنده، غشاهای گوناگونی را میسازند که اندامکهای درون یاخته را محصور میکنند.[5] با اینکه روشن است که در پیدایش اندامکها، انتقال لیپید یک فرایند اصلی است، هنوز سازوکارهای انتقال لیپیدها درک نشدهاند.[6]
نخستین پیشنهادی که بیان داشت غشای درون یاختهها دستگاهی واحد تشکیل میدهند که مواد را بین اجزای آن مبادله میکند، توسط موره[persian-alpha 1] و مولنهاور[persian-alpha 2] در سال ۱۹۷۴ ارائه شد.[7] این پیشنهاد به عنوان روشی برای توضیح چگونگی جمع شدن غشاهای چربی گوناگون در یاخته از راه جریان چربی از مکانهای ساخت چربی، ارائه شد.[8] ایده جریان چربی از راه یک دستگاه پیوسته از غشاها و وزیکولها، جایگزینی برای غشاهای گوناگون با موجودیت مستقل بود که از جابهجایی اجزای چربی آزاد، مانند اسیدهای چرب و استرولها، از طریق سیتوزول تشکیل میشوند. انتقال لیپیدها از طریق سیتوزول و جریان لیپیدها از راه یک دستگاه غشایی درونی پیوسته، فرآیندهای متقابل انحصاری نیست و هر دو ممکن است در یاختهها رخ دهند.[5] در تعریفی دقیقتر، این دستگاه مجموعهای از غشاهاست که با تشکیل یک واحد کارکردی و تکاملی، یا مستقیماً با هم پیوند دارند یا از طریق وزیکولها با هم ارتباط دارند.[9]
اجزای دستگاه
پوشش هستهای
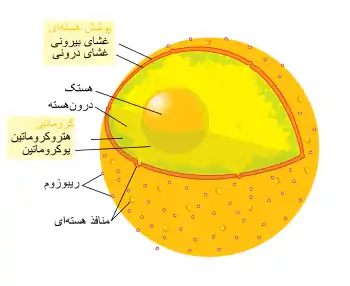
پوشش هستهای از دو لایه غشای لیپیدی دولایهٔ ساخته شدهاست و محتوای هسته را در بر میگیرد و آن را از سیتوپلاسم جدا میکند.[10][11] غشای هستهای بیرونی با غشای شبکه آندوپلاسمی زبر پیوستهاست و همانند آن روی سطح خود ریبوزومهایی چسبیده دارد. غشای هستهای بیرونی با غشای هستهای درونی پیوستهاست زیرا این دو لایه در سوراخهای کوچک زیادی به نام منفذ هستهای به هم چسبیدهاند. این منافذ که قطری نزدیک به ۱۲۰ نانومتر دارند، با اجازه دادن به عبور برخی مولکولها از غشا و اجازه ندادن به برخی دیگر، عبور و مرور از غشا را تنظیم میکنند.[12] از آنجا که منافذ هستهای در منطقهای با رفتوآمد زیاد قرار دارند، نقش مهمی در فیزیولوژی یاختهها دارند. فضای میان غشاهای خارجی و داخلی را فضای هستهای مینامند که با حفره شبکه آندوپلاسمی زبر پیوستهاست.
ساختار پوشش هستهای توسط شبکهای از رشتههای میانی (رشتههای پروتئینی) تعیین میشود. این شبکه، لایهای سازمانیافته و توریشکل به نام لامینای هسته است که به کروماتین، پروتئینهای سراسری غشایی و سایر اجزای هستهای در امتداد سطح درونی هسته متصل میشود. به نظر میرسد که لامینای هستهای به رسیدن مواد درون هسته به منافذ هستهای و از هم پاشیده شدن پوشش هستهای در هنگام میتوز و ساخت دوبارهٔ آن در پایان میتوز کمک میکند.[10]
منافذ هستهای از نظر انتخابی اجازه عبور مواد از هسته را میدهند و از هسته بسیار کارآمدتر هستند، زیرا پوشش هستهای میزان رفتوآمد قابل توجهی دارد. آرانای و زیرواحدهای ریبوزوم باید بهطور پیوسته از هسته به سیتوپلاسم منتقل شوند. هیستونها، پروتئینهای تنظیم کننده ژن، دیانای پلیمراز و آرانای پلیمراز و سایر مواد ضروری برای فعالیتهای هستهای باید از سیتوپلاسم به هسته وارد شوند. پوشش هستهای یک یاختهٔ پستاندار معمولی شامل ۳۰۰۰ تا ۴۰۰۰ مجموعهٔ منفذی است. اگر یاخته در حال ساخت دیانای باشد، هر مجموعه منفذی نیاز به انتقال حدود ۱۰۰ مولکول هیستون در دقیقه دارد. اگر یاخته به سرعت در حال رشد باشد، هر مجموعه باید حدود ۶ زیرواحد ریبوزومی بزرگ و کوچک تازه جمع شده در هر دقیقه را از هسته به سیتوزول - جایی که برای ساخت پروتئینها استفاده میشوند - منتقل کند.[13]
شبکهٔ آندوپلاسمی
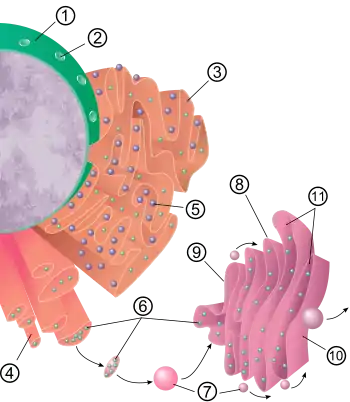
شبکهٔ آندوپلاسمی (ER) یک اندامک غشایی سازنده (سنتزکننده) و انتقالدهنده است که در یاختههای گیاهی و جانوری، در ادامهٔ پوشش هستهای در داخل سیتوپلاسم منشعب میشود.[14] بیش از نیمی از غشاهای یاختهٔ یوکاریوتی را شبکهٔ آندوپلاسمی میسازد. شبکهٔ آندوپلاسمی از کیسههای مسطح و لولههای منشعب ساخته شدهاست که تصور میشود به یکدیگر متصل باشند، به طوری که غشای شبکهٔ آندوپلاسمی یک سطح پیوسته را تشکیل میدهد که یک فضای درونی واحد را میسازند. این فضای کاملاً پیچیده لومن شبکهٔ آندوپلاسمی نامیده میشود و از آن به عنوان فضای مخزنی شبکهٔ آندوپلاسمی نیز یاد میشود. لومن حدود ده درصد از کل یاخته را اشغال میکند. غشای شبکهٔ آندوپلاسمی اجازه میدهد تا مولکولها بهطور انتخابی میان لومن و سیتوپلاسم جابهجا شوند و از آنجا که به پوشش هستهای متصل است، یک کانال میان هسته و سیتوپلاسم فراهم میکند.[15]
شبکهٔ آندوپلاسمی نقش اصلی در تولید، پردازش و انتقال ترکیبهای بیوشیمیایی برای استفاده در درون و بیرون یاخته را دارد. غشای آن جایگاه ساخت پروتئینها و چربیهای تراغشایی برای بیشتر اندامکهای یاخته از جمله خود شبکهٔ آندوپلاسمی، دستگاه گلژی، لیزوزوم، اندوزوم، میتوکندری، پراکسیزوم، وزیکولهای ترشحی و غشای پلاسما است. افزون بر این، تقریباً همهٔ پروتئینهایی که قرار است از یاخته خارج شوند، به علاوهٔ آنهایی که برای لومن شبکهٔ آندوپلاسمی، دستگاه گلژی یا لیزوزومها اختصاص دارند، در اصل به لومن شبکهٔ آندوپلاسمی منتقل میشوند. در نتیجه، بسیاری از پروتئینهای موجود در فضای کیسهای لومن شبکهٔ آندوپلاسمی، حضوری موقتی دارند زیرا در مسیر خود به مکانهای دیگر منتقل میشوند. پروتئینهای دیگر، بهطور پیوسته در لومن باقی میمانند و به عنوان پروتئینهای ساکن شبکهٔ آندوپلاسمی شناخته میشوند. این پروتئینهای ویژه دارای یک سیگنال ویژهٔ نگهداری هستند که از توالی ویژهای از آمینواسیدها تشکیل شدهاست که باعث نگهداری آنها توسط اندامک میشود. یک نمونه از پروتئینهای مهم ساکن شبکهٔ آندوپلاسمی، پروتئین شپرون معروف به پروتئین اتصالدهنده ایمونوگلوبولین (BiP) است که پروتئینهای دیگری را که بهطور نامناسب ساخته یا پردازش شدهاند، شناسایی کرده و آنها را از ارسال به مقصد نهایی بازمیدارد.[16]
شبکهٔ آندوپلاسمی در مرتبسازی پروتئینها بر پایهٔ ترجمه نقش دارد. پلیپپتیدی که دارای توالی سیگنالی شبکهٔ آندوپلاسمی است، توسط یک پروتئین تشخیص سیگنال که تولید پروتئین را متوقف میکند، شناسایی میشود. ذره شناسایی سیگنال (SRP) پلیپپتید را به غشای شبکهٔ آندوپلاسمی منتقل میکند تا در آنجا، از طریق منفذهای غشایی رها شده و ترجمه از سر گرفته شود.[17]
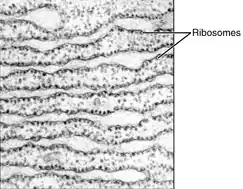
دو بخش متمایز اما بههمپیوسته از شبکهٔ آندوپلاسمی وجود دارد که از لحاظ ساختار و کارکرد گوناگون هستند: شبکهٔ آندوپلاسمی صاف و شبکهٔ آندوپلاسمی زبر. دلیل نامگذاری شبکهٔ آندوپلاسمی زبر وجود ریبوزومها در سطح سیتوپلاسمی آن است. این نوع شبکه آندوپلاسمی هنگام مشاهده با میکروسکوپ الکترونی، ظاهری پر از دستانداز دارد. شبکهٔ آندوپلاسمی صاف از آنجا که سطح سیتوپلاسمی آن فاقد ریبوزوم است، صاف به نظر میرسد.[18]
کارکرد شبکهٔ آندوپلاسمی صاف
در بیشتر یاختهها، نواحی کاملاً صاف شبکهٔ آندوپلاسمی، کمیاباند و این شبکهها بیشتر بهصورت نیمهصاف -نیمهزبر هستند. به شبکهٔ آندوپلاسمی صاف گاه شبکهٔ آندوپلاسمی انتقالی نیز گفته میشود، زیرا دارای جایگاههای خروجی است که از آنها وزیکولهای حامل پروتئینها و لیپیدهای تازه ساخته شده برای انتقال به دستگاه گلژی خارج میشوند. با این حال، در برخی یاختههای ویژه، شبکهٔ آندوپلاسمی صاف فراوان است و کارکردهای بیشتری دارد. کارکردهای شبکهٔ آندوپلاسمی این یاختههای ویژه، در فرآیندهای سوختوسازی گوناگون از جمله ساخت لیپیدها، سوختوساز کربوهیدراتها و سمزدایی از داروها و سموم است.[15][18]
آنزیمهای شبکهٔ آندوپلاسمی صاف برای ساخت لیپیدها از جمله روغنها، فسفولیپیدها و استروئیدها حیاتی هستند. هورمونهای جنسی مهرهداران و هورمونهای استروئیدی ترشحشده از غدد فوق کلیوی از جمله استروئیدهای ساخته شده توسط شبکهٔ آندوپلاسمی صاف در یاختههای جانوری هستند. یاختههایی که این هورمونها را میسازند، سرشار از شبکهٔ آندوپلاسمی صاف هستند.[15][18]
یاختههای کبد نمونه دیگری از یاختههای ویژهای هستند که دارای شبکهٔ آندوپلاسمی صاف فراوانی هستند. این یاختهها نمونهای از نقش شبکهٔ آندوپلاسمی صاف در سوختوساز کربوهیدرات را نشان میدهند. یاختههای کبدی کربوهیدراتها را به صورت گلیکوژن ذخیره میکنند. تجزیه گلیکوژن در نهایت منجر به آزاد شدن گلوکز از یاختههای کبدی میشود که در تنظیم غلظت قند خون مهم است. با این حال، محصول اصلی تجزیه گلیکوژن گلوکز-۱-فسفات است. این ماده به گلوکز-۶-فسفات تبدیل میشود و سپس یک آنزیم شبکهٔ آندوپلاسمی صاف یاخته کبدی فسفات را از گلوکز خارج میکند، به طوری که پس از آن میتواند یاخته را ترک کند.[15][18]
آنزیمهای شبکهٔ آندوپلاسمی صاف همچنین میتوانند به سمزدایی داروها و سموم کمک کنند. سم زدایی معمولاً با افزودن یک گروه هیدروکسیل به دارو باعث حل شدن بیشتر دارو و در نتیجه پاک سازی بدن میشود. یک واکنش سم زدایی بهطور گسترده توسط خانوادهٔ آنزیمهای سیتوکروم پی ۴۵۰ انجام میشود که داروها یا متابولیتهای محلول در آبی را کاتالیز میکند تا سطوح سمی در غشای یاخته تجمع نیابند.[15][18]
کارکرد ویژهٔ دیگری از شبکهٔ آندوپلاسمی صاف در یاختههای ماهیچهای دیدهمیشود. غشا شبکهٔ آندوپلاسمی یونهای کلسیم را از سیتوزول به فضای حفرهای پمپ میکند. هنگامی که یک یاخته ماهیچهای تحت فشار عصبی تحریک میشود، کلسیم از طریق غشای شبکهٔ آندوپلاسمی به درون سیتوزول بر میگردد و انقباض یاخته ماهیچهای را ایجاد میکند.[15][18]
کارکرد شبکهٔ آندوپلاسمی زبر
بسیاری از انواع یاختهها، قادرند پروتئینهای ساخته شده توسط ریبوزومهای متصل به شبکهٔ آندوپلاسمی زبر را برونریزی کنند. ریبوزومها از آمینواسیدها، واحدهایی پروتئینی میسازند که برای تنظیم و تعدیل بیشتر، به شبکهٔ آندوپلاسمی زبر منتقل میشوند. این پروتئینها ممکن است یا پروتئینهای تراغشایی باشند که در غشای شبکهٔ آندوپلاسمی قرار میگیرند یا پروتئینهای محلول در آب، که میتوانند با عبور از غشا به لومن وارد شوند. پروتئینهایی که به داخل شبکهٔ آندوپلاسمی میرسند، از چینخوردگیهای سهبعدی درستی در ساختار خود برخوردار خواهند بود. مواد شیمیایی مانند کربوهیدراتها یا قندها به آن افزوده میشود، سپس شبکهٔ آندوپلاسمی، پروتئینهای کاملشده را که «پروتئینهای ترشحی» نامیده میشون، به مناطقی از یاخته که مورد نیاز است منتقل میکند یا آنکه برای پردازش و اصلاح بیشتر به دستگاه گلژی ارسال میکند.[15][18]
به محض تشکیل پروتئینهای ترشحی، غشای شبکهٔ آندوپلاسمی آنها را از پروتئینهای باقیمانده در سیتوزول جدا میکند. پروتئینهای ترشحی با وزیکولهای غشایی حبابمانند از شبکهٔ آندوپلاسمی انتقالی بیرون میآیند. به این وزیکولهایِ در حالِ انتقال به بخشهایِ دیگری از یاخته، وزیکولهای ناقل گفته میشود.[15][18] پروتئینهای انتقال لیپید موجود در محل تماس غشایی، راهکاری جایگزین برای انتقال لیپیدها و پروتئینها به خارج از شبکهٔ آندوپلاسمی هستند. محل تماس غشایی، مکانی است که شبکهٔ آندوپلاسمی با غشای دیگر اندامکها مانند غشای پلاسما، گلژی یا لیزوزوم ارتباط نزدیک و پایداری دارد.[19]
شبکهٔ آندوپلاسمی زبر، افزون بر ساخت پروتئینهای ترشحی، سبب ایجاد غشایی میشود که در اثر افزودن پروتئینها و فسفولیپیدها در محل رشد میکنند. از آنجا که پلیپپتیدهای در نظر گرفتهشده به عنوان پروتئینهای غشایی از ریبوزومها رشد میکنند، آنها در غشای شبکهٔ آندوپلاسمی قرار میگیرند و توسط بخشهای آبگریز در آنجا نگهداری میشوند. شبکهٔ آندوپلاسمی زبر نیز فسفولیپیدهای غشایی خود را تولید میکند. آنزیمهای ساخته شده در غشای شبکهٔ آندوپلاسمی فسفولیپیدها را میسازند. غشای شبکهٔ آندوپلاسمی قابلیت گسترش دارد و میتواند توسط وزیکولهای ناقل به سایر اجزای دستگاه غشایی درونی منتقل شود.[15][18]
دستگاه گلژی
دستگاه گلژی مجموعهای از چند کیسه است که در آن مولکولها برای تحویل به سایر اجزای یاخته یا ترشح از یاخته، بستهبندی میشوند.[20] این دستگاه از کیسههای جداگانهای به نام سیسترنا تشکیل شدهاست. شکل آن شبیه چند پنکیک است که روی هم قرار گرفتهاند. شمار این دستهها با کارکرد ویژه یاخته متفاوت است. یاخته با استفاده از دستگاه گلژی، اصلاح بیشتری روی پروتئینها انجام میدهند. بخشی از دستگاه گلژی که وزیکولها را از شبکه آندوپلاسمی دریافت میکند، به عنوان «رویهٔ سیس» شناخته میشود و معمولاً نزدیک شبکه آندوپلاسمی است. انتهای مخالف دستگاه گلژی «رویهٔ ترانس» نامیده میشود که همان جایی است که ترکیبات اصلاحشده، دستگاه گلژی را ترک میکنند. رویهٔ ترانس معمولاً رو به غشای پلاسمایی است که دستگاه گلژی بیشتر مواد را به آنجا میفرستد.[21]
وزیکولهای فرستادهشده توسط شبکهٔ آندوپلاسمی حاوی پروتئینها، در دستگاه گلژی تغییرات بیشتری مییابند و سپس برای ترشح از یاخته یا انتقال به سایر قسمتهای یاخته آماده میشوند. اتفاقهای گوناگونی ممکن است برای پروتئینها طی عبور از طریق فضای آنزیمی دستگاه گلژی رخ دهند. اصلاح و ساخت بخشهای کربوهیدرات گلیکوپروتئینها در پردازش پروتئین معمول است. دستگاه گلژی مونومرهای قند را حذف و جایگزین و الیگوساکاریدهای زیادی را تولید میکند. گلژی افزون بر اصلاح پروتئینها، خود درشتمولکولها را نیز تولید میکند. گلژی در یاختههای گیاهی پکتین و سایر پلیساکاریدهای مورد نیاز ساختار گیاه را تولید میکند.[22]
دستگاه گلژی پس از اتمام فرایند اصلاح، محصولات فرآوریشده را طبقهبندی کرده و به قسمتهای مختلف یاخته میفرستد. آنزیمهای گلژی برای کمک به این موارد، برچسبها یا برچسبهای شناسایی مولکولی را اضافه میکنند. بعد از اینکه همه چیز مرتب شد، دستگاه گلژی محصولهای خود را با وزیکولهایی که از غشای آن جوانه میزنند، از رویهٔ ترانس خود ارسال میکند.[23]
واکوئل
واکوئلها، مانند وزیکولها، کیسههای غشایی درون یاخته و بزرگتر از وزیکولها هستند. آنها هم در یاختههای گیاهی و هم در جانوران یافت میشوند (اگرچه در یاختههای گیاهی بسیار بزرگتر هستند) و مسئول حفظ شکل و ساختار یاخته و همچنین ذخیره مواد زائد هستند.[24] کارکرد واکوئلها در گیاهان و جانوران متفاوت است.
در یاختههای گیاهی، واکوئلها، ۳۰ تا ۹۰ درصد از کل یاخته را پوشش میدهند.[25] بیشتر یاختههای گیاهی بالغ دارای یک واکوئل مرکزی بزرگ هستند که توسط غشایی به نام تونوپلاست احاطه شدهاست. واکوئلهای یاختههای گیاهی به عنوان محفظه ذخیره مواد مغذی و مواد زائد یاخته عمل میکنند. به محلولی که این مولکولها در آن ذخیره شدهاند، شیره یاخته گفته میشود. رنگدانههایی که یاخته را رنگ میکنند، گاهی در شیره یاخته قرار دارند. واکوئلها همچنین میتوانند اندازه یاخته را افزایش دهند، که با افزودن آب کشیده و بزرگ میشوند و آماسیدگی یا تورژسانس (فشار اسمزی که باعث میشود دیواره یاخته متلاشی نشود) را کنترل میکنند. مانند لیزوزوم یاختههای جانوری، واکوئلها دارای پیاچ اسیدی و آنزیمهای هیدرولیتیک هستند. پیاچ واکوئلها آنها را توانا میکند تا مراحل هموستاتیک را در یاخته انجام دهند. برای نمونه، هنگامی که پیاچ در محیط یاخته کاهش مییابد، یونهای +H که در سیتوزول قرار میگیرند میتوانند به واکوئل منتقل شوند تا پیاچ سیتوزول ثابت بماند.[26]
واکوئلها در جانوران در فرآیندهای اگزوسیتوز (برونرانی) و آندوسیتوز (درونبری) خدمت میکنند. در آندوسیتوز مواد از فضای میانیاختهای وارد یاخته شده و در اگزوسیتوز مواد از یاخته به فضای برونیاختهای منتقل میشوند. غشای پلاسما موادی که باید جابهجا شوند را احاطه و سپس به واکوئل منتقل میکند. آندوسیتوز دو نوع است، فاگوسیتوز (بیگانهخواری) و پینوسیتوز (قطرهخواری). در فاگوسیتوز، یاختهها ذرات بزرگی مانند باکتریها را میبلعند اما در پینوسیتوز یاخته مواد مایع را به درون میآورند.[27]
وزیکول
وزیکولها واحدهای کوچک غشایی و ناقل هستند که میتوانند مولکولها را ذخیره و میان بخشهای گوناگون جابهجا کنند.[28] بیشتر وزیکولها غشاهای ساخته شده در شبکه آندوپلاسمی را به دستگاه گلژی و سپس از دستگاه گلژی به مکانهای مختلف منتقل میکنند.[29]
انواع گوناگونی از وزیکولها وجود دارند که هر یک پیکربندی پروتئینی متفاوت دارند. بیشتر آنها از مناطق ویژهای از غشا ساخته میشوند. هنگامی که وزیکول از غشا جوانه میزند، دارای پروتئینهای ویژهای در سطح سیتوزولی آن است. هر غشایی که وزیکول به سمت آن میرود در سطح سیتوزولی خود نشانگری دارد. این نشانگر با پروتئینهای موجود در وزیکول در حال حرکت به سمت غشا مطابقت دارد. هنگامی که وزیکول غشا را پیدا میکند، به آن اضافه میشود و سطح آن غشا را افرایش میدهد.[30]
سه نوع وزیکول شناخته شده وجود دارد: وزیکولهای پوشیدهشده با کلاترین، پوشیدهشده با COPI و پوشیدهشده با COPII. هر یک از این وزیکولها کارکردهای گوناگونی در یاخته دارند. برای نمونه، وزیکولهای پوشیدهشده با کلاترین مواد را میان دستگاه گلژی و غشای پلاسما جابهجا میکنند. از وزیکولهای پوشیدهشده با COPI و COPII بیشتر برای جابهجایی میان شبکه آندوپلاسمی و دستگاه گلژی استفاده میشود.[30]
لیزوزوم
لیزوزومها اندامکهایی دارای آنزیمهای هیدرولاز هستند که برای گوارش درونیاختهای استفاده میشوند. کارهای اصلی لیزوزوم پردازش مولکولهای جذبشده توسط یاخته و بازیافت قطعههای فرسوده یاخته است. آنزیمهای داخل لیزوزومها هیدرولاز اسیدها هستند که برای کارکرد بهینه به محیط اسیدی نیاز دارند. لیزوزومهای اندامکها با ثابت نگهداشتن اندازه پیاچ در ۵٫۰، چنین محیطی را فراهم میکنند.[31] اگر یک لیزوزوم پاره شود، آنزیمهای آزادشده به دلیل پیاچ خنثی سیتوزول برای مدت زیادی فعال نخواهند بود. با این حال، اگر لیزوزومهای زیادی پاره شوند، یاخته ممکن است در اثر گوارش خودش از بین برود.
لیزوزومها با ترکیب شدن با واکوئل و آزادسازی آنزیمهای خود در واکوئل، گوارش درونیاختهای را طی فرآیندی به نام فاگوسیتوز انجام میدهند. قندها، آمینواسیدها و سایر مونومرها از طریق این فرایند به سیتوزول منتقل میشوند و به عناصر غذایی یاخته تبدیل میشوند. لیزوزومها همچنین در فرآیندی به نام خودخواری از آنزیمهای هیدرولیتیک خود برای بازیافت اندامهای فرسوده یاخته استفاده میکنند. لیزوزوم اندامک فرسوده را میبلعد و آنزیمهای آن اندامک را تجزیه میکنند. سپس مونومرهای آلی حاصل برای استفاده دوباره به سیتوزول بازگردانده میشوند. آخرین عملکرد لیزوزوم گوارش خود یاخته از طریق خودکافت است.[32]
اشپیتزنکاپه
اشپیتزنکاپه بخشی از دستگاه غشایی درونی است که فقط در قارچها یافت میشود و با رشد نوک هیف مرتبط است. این یک جسم تاریک فاز است که از تجمع وزیکولهای متصل به غشا دارای اجزای دیوارهٔ یاختهای تشکیل شدهاست و به عنوان نقطهای برای ساخت و آزادسازی چنین اجزای میانی بین گلژی و غشای یاخته کار میکند. اشپیتزنکاپه متحرک است و با حرکت رو به جلو رشد نوک هیف جدیدی را آغاز میکند.[33]
غشای یاختهای
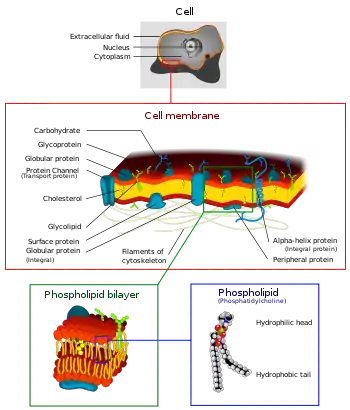
غشای پلاسما غشایی ساخته شده از دو لایه فسفولیپید است که درون یاخته را از محیط جدا میکند و انتقال مولکولها و سیگنالها را به داخل و خارج یاخته تنظیم میکند. پروتئینهای جاسازی شده در غشا، کارهای غشا را انجام میدهند. غشای پلاسما یک ساختار ثابت یا سفت و سخت نیست و مولکولهای سازندهٔ آن توانایی حرکت جانبی دارند. این حرکت و اجزای متعدد غشا باعث شدهاند که از آن به عنوان موزاییک سیال یاد میشود. مولکولهای کوچکتری مانند کربن دیاکسید، آب و اکسیژن میتوانند با انتشار یا اسمز، آزادانه از غشای پلاسما عبور کنند. مولکولهای بزرگتر مورد نیاز یاخته توسط پروتئینها از طریق انتقال فعال از غشا عبور میکنند.[34]
غشای پلاسمای یاخته کارکردهای پرشماری دارد. این موارد شامل انتقال مواد مغذی به داخل یاخته، اجازه خروج مواد زائد، جلوگیری از ورود مواد به یاخته، جلوگیری از خروج مواد مورد نیاز از یاخته، حفظ پیاچ و فشار اسمزی سیتوزول است. پروتئینهای انتقالی که فقط به برخی از مواد اجازه عبور میدهند، برای این کارکردها استفاده میشوند. این پروتئینها با دریافت انرژی پس از هیدرولیز ATP، مواد را در خلاف شیب غلظت پمپاژ میکنند.[34]
غشای پلاسما افزون بر این کارکردهای کلی نقش ویژهتری در جانداران چند یاختهای دارد. گلیکوپروتئینهای موجود در غشا به یاخته در شناسایی یاختههای دیگر برای تبادل متابولیتها و تشکیل بافتها، کمک میکنند. دیگر پروتئینهای غشای پلاسما امکان اتصال به اسکلت یاختهای و ماتریکس برونیاختهای را فراهم میکنند؛ کارکردی که شکل یاخته را حفظ کرده و محل پروتئینهای غشایی را ثابت میکند. آنزیمهایی که واکنشها را کاتالیز میکنند نیز در غشای پلاسما یافت میشوند. پروتئینهای گیرنده روی غشا شکلی دارند که با یک پیامرسان شیمیایی مطابقت دارد و در نتیجه پاسخهای یاختهای گوناگونی ایجاد میشود.[35]
ارتباط میان اجزا
اندامکهای دستگاه غشایی درونی یا از راه تماس مستقیم یا با انتقال بخشهای غشایی به صورت وزیکول، با یکدیگر ارتباط دارند. با وجود این روابط، غشاهای گوناگون از نظر ساختار و کارکرد یکسان نیستند. ضخامت، ترکیب مولکولی و رفتار سوخت وسازی یک غشا ثابت نیستند و ممکن است چندین بار در طول زندگی غشا اصلاح شوند. یکی از ویژگیهای یکسان غشاها، دو لایه چربی است که پروتئینها به هر دو طرف متصل شدهاند یا از آنها عبور میکنند.[36]
تکامل
خاستگاه دستگاه غشایی درونی به خاستگاه خود یوکاریوتها و خاستگاه یوکاریوتیها به خاستگاه درونهمزیستانهٔ میتوکندری مرتبط است. مدلهای زیادی برای توضیح خاستگاه دستگاه غشایی درونی ارائه شدهاست (بررسی شده در[37]). جدیدترین پیشنهاد نشان میدهد که دستگاه غشایی درونی، از وزیکولهای ترشحشده غشای خارجی میتوکندری (OMV)، تکامل یافتهاست.[38] این مدلِ مبتنی بر OMV برای توضیحِ خاستگاه دستگاه غشایی درونی، مدلی است که در حال حاضر پذیرفته شدهاست و نیاز چندانی به تازهاندیشی برای یافتن منشأ یوکاریوتها ندارد و بسیاری از پیوندهای میتوکندری را با دیگر بخشهای یاخته توضیح میدهد.[39]
در پروکاریوتها
در پروکاریوتها دستگاه غشایی درونی نادر است، اگرچه در بسیاری از باکتریهای فتوسنتزکننده غشای پلاسما بسیار چین خورده و بیشتر سیتوپلاسم یاخته با لایههای غشای جمعکننده نور پر شدهاست.[40] این غشاهای جمعکننده نور حتی ممکن است در باکتریهای گوگردی سبز ساختارهای محصور به نام کلروزوم ایجاد کنند.[41]
پانویس
- D. James Morré
- Hilton H. Mollenhauer
منابع
- Zinser E, Sperka-Gottlieb CD, Fasch EV, Kohlwein SD, Paltauf F, Daum G (March 1991). "Phospholipid synthesis and lipid composition of subcellular membranes in the unicellular eukaryote Saccharomyces cerevisiae". Journal of Bacteriology. 173 (6): 2026–34. doi:10.1128/jb.173.6.2026-2034.1991. PMC 207737. PMID 2002005.
- Czabany T, Athenstaedt K, Daum G (March 2007). "Synthesis, storage and degradation of neutral lipids in yeast". Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids. 1771 (3): 299–309. doi:10.1016/j.bbalip.2006.07.001. PMID 16916618.
- Futerman AH (December 2006). "Intracellular trafficking of sphingolipids: relationship to biosynthesis". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1758 (12): 1885–92. doi:10.1016/j.bbamem.2006.08.004. PMID 16996025.
- Wanders RJ, Waterham HR (2006). "Biochemistry of mammalian peroxisomes revisited". Annual Review of Biochemistry. 75: 295–332. doi:10.1146/annurev.biochem.74.082803.133329. PMID 16756494.
- Voelker DR (December 1991). "Organelle biogenesis and intracellular lipid transport in eukaryotes". Microbiological Reviews. 55 (4): 543–60. doi:10.1128/MMBR.55.4.543-560.1991. PMC 372837. PMID 1779926.
- Voelker DR (July 2005). "Bridging gaps in phospholipid transport". Trends in Biochemical Sciences. 30 (7): 396–404. doi:10.1016/j.tibs.2005.05.008. PMID 15951180.
- Morré DJ, Mollenhauer HH (1974). "The endomembrane concept: a functional integration of endoplasmic reticulum and Golgi apparatus.". In Robards AW. Dynamic Aspects of Plant infrastructure. London, New York: McGraw-Hill. pp. 84–137.
- Morre DJ (1975). "Membrane Biogenesis". Annual Review of Plant Physiology. 26 (1): 441–481. doi:10.1146/annurev.pp.26.060175.002301.
- Smith AL (1997). Oxford dictionary of biochemistry and molecular biology. Oxford [Oxfordshire]: Oxford University Press. pp. 206. ISBN 978-0-19-854768-6.
- Davidson, Michael (2005). "The Nuclear Envelope". Molecular Expressions. Florida State University. Retrieved 2008-12-09.
- Childs, Gwen V. (2003). "Nuclear Envelope". UTMB. Archived from the original on June 20, 2006. Retrieved 2008-09-28.
- Cooper, Geoffrey (2000). "The Nuclear Envelope and Traffic between the Nucleus and Cytoplasm". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Alberts, Walter; et al. (2002). "Nuclear Pore Complexes Perforate the Nuclear Envelope". Molecular Biology of the Cell 4th edition. Garland Science. Retrieved 2008-12-09.
- Davidson, Michael (2005). "The Endoplasmic Reticulum". Molecular Expressions. Florida State University. Retrieved 2008-12-09.
- Cooper, Geoffrey (2000). "The Endoplasmic Reticulum". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D (June 2000). "Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response". Nature Cell Biology. 2 (6): 326–32. doi:10.1038/35014014. PMID 10854322. S2CID 22684712.
- Biology. McGraw Hill education. 2011. pp. 89.
- Alberts, Walter; et al. (2002). "Membrane-bound Ribosomes Define the Rough ER". Molecular Biology of the Cell 4th edition. Garland Science. Retrieved 2008-12-09.
- Levine T, Loewen C (August 2006). "Inter-organelle membrane contact sites: through a glass, darkly". Current Opinion in Cell Biology. 18 (4): 371–8. doi:10.1016/j.ceb.2006.06.011. PMID 16806880.
- Graham, Todd R. (2000). Eurekah Bioscience Collection Cell Biology. University of New South Wales and Landes Bioscience. ISBN 978-0-7334-2108-2.
- Rothman JE (September 1981). "The golgi apparatus: two organelles in tandem". Science. 213 (4513): 1212–9. Bibcode:1981Sci...213.1212R. doi:10.1126/science.7268428. PMID 7268428.
- Alberts, Walter; et al. (2002). "Transport from the ER through the Golgi Apparatus". Molecular Biology of the Cell 4th edition. Garland Science. Retrieved 2008-12-09.
- Cooper, Geoffrey (2000). "The Golgi Apparatus". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Lodish, Harvey; et al. (2000). "Section 5.4 Organelles of the Eukaryotic Cell". Molecular Cell Biology. W. H. Freeman and Company. Retrieved 2008-12-09.
- Alberts, Walter; et al. (2002). "Plant and Fungal Vacuoles Are Remarkably Versatile Lysosomes". Molecular Biology of the Cell 4th edition. Garland Science. Retrieved 2008-12-09.
- Lodish, Harvey; et al. (2000). "Plant Vacuoles Store Small Molecules and Enable the Cell to Elongate Rapidly". Molecular Cell Biology. W. H. Freeman and Company. Retrieved 2008-12-09.
- Cooper, Geoffrey (2000). "Endocytosis". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Cooper, Geoffrey (2000). "The Mechanism of Vesicular Transport". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Lodish, Harvey; et al. (2000). "Section 17.10 Molecular Mechanisms of Vesicular Traffic". Molecular Cell Biology. W. H. Freeman and Company. Retrieved 2008-12-09.
- Alberts, Walter; et al. (2002). "The Molecular Mechanisms of Membrane Transport and the Maintenance of Compartmental Diversity". Molecular Biology of the Cell 4th edition. Garland Science. Retrieved 2008-12-09.
- Alberts, Walter; et al. (2002). "Transport from the Trans Golgi Network to Lysosomes". Molecular Biology of the Cell 4th edition. Garland Science. Retrieved 2008-12-09.
- Cooper, Geoffrey (2000). "Lysosomes". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Steinberg G (March 2007). "Hyphal growth: a tale of motors, lipids, and the Spitzenkörper". Eukaryotic Cell. 6 (3): 351–60. doi:10.1128/EC.00381-06. PMC 1828937. PMID 17259546.
- Cooper, Geoffrey (2000). "Structure of the Plasma Membrane". The Cell: A Molecular Approach. Sinauer Associates, Inc. Retrieved 2008-12-09.
- Lodish, Harvey; et al. (2000). "Section 5.3. Biomembranes: Structural Organization and Basic Functions". Molecular Cell Biology. W. H. Freeman and Company. Retrieved 2008-12-09.
- Campbell, Neil A.; Reece, Jane B. (2002). Biology (6th ed.). Benjamin Cummings. ISBN 978-0-8053-6624-2.
- Martin WF, Garg S, Zimorski V (September 2015). "Endosymbiotic theories for eukaryote origin". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1678): 20140330. doi:10.1098/rstb.2014.0330. PMC 4571569. PMID 26323761.
- Gould SB, Garg SG, Martin WF (July 2016). "Bacterial Vesicle Secretion and the Evolutionary Origin of the Eukaryotic Endomembrane System". Trends in Microbiology. 24 (7): 525–534. doi:10.1016/j.tim.2016.03.005. PMID 27040918.
- Murley A, Nunnari J (March 2016). "The Emerging Network of Mitochondria-Organelle Contacts". Molecular Cell. 61 (5): 648–653. doi:10.1016/j.molcel.2016.01.031. PMC 5554544. PMID 26942669.
- Bryant DA, Frigaard NU (November 2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends in Microbiology. 14 (11): 488–96. doi:10.1016/j.tim.2006.09.001. PMID 16997562.
- Psencík J, Ikonen TP, Laurinmäki P, Merckel MC, Butcher SJ, Serimaa RE, Tuma R (August 2004). "Lamellar organization of pigments in chlorosomes, the light harvesting complexes of green photosynthetic bacteria". Biophysical Journal. 87 (2): 1165–72. Bibcode:2004BpJ....87.1165P. doi:10.1529/biophysj.104.040956. PMC 1304455. PMID 15298919. Archived from the original on 10 May 2020. Retrieved 27 April 2021.
