لیپید اتری
از نظر کلی شیمی آلی، یک لیپید اتر به معنای پل اتر بین یک گروه آلکیل (یک لیپید) و یک گروه آلکیل یا آریل نامشخص (نه لزوماً گلیسرول) است. اگر گلیسرول درگیر باشد، این ترکیب گلیسیریل اتر نامیده میشود که ممکن است به شکل یک آلکیل گلیسرول یا یک آلکیل آسیل گلیسرول یا در ترکیب با یک گروه فسفاتید، یک فسفولیپید باشد.
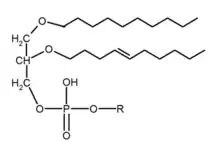
از نظر بیوشیمی، یک لیپید اتر معمولاً به گلیسروفسفولیپیدهای گوناگون اشاره میکند که به آنها فسفولیپید نیز گفته میشود که در آن موقعیت sn-1 بنیان گلیسرول دارای یک لیپید متصلشده توسط یک پیوند اتر و یک لیپید متصل به موقعیت sn-2 از طریق یک گروه آسیل است. این در تضاد با گلیسروفسفولیپیدهای معمولتر، ۱٬۲-دیاسیل-اسان-گلیسرول (DAG) است که در آن بنیان گلیسرول موقعیتهای sn-1 و sn-2 دارای زنجیرههای آسیل متصلشده توسط پیوندهای استر هستند.[1][2] لیپید اتر همچنین ممکن است به آلکیلگلیسرولها، مانند الکلهای کیمیل (۱۶:۰)، باتیل (۱۸:۰)، و سلاچیل (۱۸:۱ n-۹)، با چربی متصل به اتر بر روی موقعیت sn-1، و دو موقعیت دیگر بر روی بنیان گلیسرول خالی، اشاره داشته باشد.[3]
گونهها
دو گونه لیپید اتر وجود دارد: پلاسمانیل-فسفولیپید و پلاسمنیل-فسفولیپید. پلاسمانیل-فسفولیپیدها در موقعیت sn-1 به یک گروه آلکیل پیوند اتر دارند. پلاسمنیل-فسفولیپیدها دارای یک پیوند اتر در موقعیت sn-1 به یک گروه آلکنیل، ۱–۰-آلک-۱‘-انیل-۲-آسیل-اسان-گلیسرول (AAG) هستند.[2] گونه اخیر پلاسمالوژن نامیده میشود.[4]
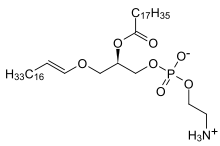
فاکتور فعالکننده پلاکتی (PAF) یک لیپید اتر است که به جای یک زنجیره آسیل در موقعیت دوم (SN-2) دارای یک گروه استیل است.
زیستساخت
تشکیل پیوند اتر در پستانداران به دو آنزیم دیهیدروکسیاستونفسفات آسیلترانسفراز (DHAPAT) و آلکیلدیهیدروکسیاستونفسفات سنتاز (ADAPS) نیاز دارد که در پراکسیزوم قرار دارند.[5] بر این اساس، نقایص پراکسیزومی اغلب منجر به اختلال در ساخت لیپید اتر میشود.
مونوالکیلگلیسرول اترها (MAGE) نیز از ۲-استیل MAGE (پیشسازهای PAF) توسط KIAA1363 ساخته میشوند.
کارکرد
ساختاری
پلاسمالوژنها و برخی لیپیدهای ۱-O-آلکیل در همه جا و گاهی بخشهای فراوانی از غشاهای یاختهای در پستانداران و باکتریهای بیهوازی هستند.[6] در باستانیان لیپید اترها مهمترین چربیهای قطبی موجود در پوشش یاخته هستند و فراوانی آنها یکی از مهمترین ویژگیهایی است که این گروه از پروکاریوتها را از باکتریها جدا میکند. در این سلولها، دیفیتانیلگلیسرولیپیدها یا تترآترهای ماکروسایکل دو قطبی میتوانند «لایههای دوتایی» کووالانسی تشکیل دهند.[7]
پیامرسان ثانویه
تفاوت میان کاتابولیسم گلیسروفسفولیپید اترها توسط فسفولیپازهای ویژه ممکن است در تولید سامانههای پیامرسان ثانویه لیپیدی مانند پروستاگلاندینها و اسید آراشیدونیک که در انتقال پیام مهم هستند، نقش داشته باشد.[8] لیپید اترها همچنین میتوانند مستقیماً در سیگنالدهی یاخته نقش داشته باشند، چرا که فاکتور فعالکننده پلاکتی یک مولکول سیگنالدهنده لیپید اتر است که در کارکرد گلبولهای سفید در دستگاه ایمنی پستانداران نقش دارد.[9]
آنتیاکسیدان
یکی دیگر از کارکردهای احتمالی لیپید اترهای پلاسمالوژن، ویژگی آنتیاکسیدانی آنها است، زیرا در برابر استرس اکسیداتیو در کشت یاخته اثرهای حافظتی نشان میدهند و بنابراین این لیپیدها ممکن است در متابولیسم لیپوپروتئین سرم نقش داشته باشند.[10] این فعالیت آنتیاکسیدانی ناشی از پیوند دوگانه اتر انول است که توسط گونههای اکسیژن فعال مورد هدف قرار میگیرد.[11]
آنالوگهای لیپید اتر مصنوعی
آنالوگهای لیپید اتر مصنوعی دارای ویژگیهای سیتواستاتیک و سیتوتوکسی هستند که احتمالاً با برهم زدن ساختار غشایی به عنوان بازدارنده آنزیم آنزیمهای درون مسیرهای انتقال سیگنال، مانند پروتئین کیناز سی و فسفولیپاز سی عمل میکنند.
میلتفوسین آنالوگ لیپید اتر سمی به تازگی به عنوان درمانی خوراکی برای بیماری گرمسیری لیشمانیاز که ناشی از انگلی پروتوزوئا به نام لیشمانیا است که لیپید اتر بسیاری در غشاهای آن دیده میشود، معرفی شدهاست.[12]
منابع
- Dean JM, Lodhi IJ (February 2018). "Structural and functional roles of ether lipids". Protein & Cell. 9 (2): 196–206. doi:10.1007/s13238-017-0423-5. PMC 5818364. PMID 28523433.
- Ford DA, Gross RW (July 1990). "Differential metabolism of diradyl glycerol molecular subclasses and molecular species by rabbit brain diglyceride kinase". The Journal of Biological Chemistry. 265 (21): 12280–6. PMID 2165056. S2CID 1042240.
- Christie, William. "Ether lipids - glyceryl ethers, plasmalogens, aldehydes, structure, biochemistry, composition and analysis". www.lipidhome.co.uk.
- Watson, Ronald Ross; De Meester, Fabien, eds. (2014). Omega 3 fatty acids in brain and neurological health. Elsevier Academic Press. doi:10.1016/C2012-0-06006-1. ISBN 978-0-12-410527-0.
- Hajra AK (1995). "Glycerolipid biosynthesis in peroxisomes (microbodies)". Progress in Lipid Research. 34 (4): 343–64. doi:10.1016/0163-7827(95)00013-5. PMID 8685243.
- Paltauf F (December 1994). "Ether lipids in biomembranes". Chemistry and Physics of Lipids. 74 (2): 101–39. doi:10.1016/0009-3084(94)90054-X. PMID 7859340.
- Koga Y, Morii H (November 2005). "Recent advances in structural research on ether lipids from archaea including comparative and physiological aspects". Bioscience, Biotechnology, and Biochemistry. 69 (11): 2019–34. doi:10.1271/bbb.69.2019. PMID 16306681.
- Spector AA, Yorek MA (September 1985). "Membrane lipid composition and cellular function". Journal of Lipid Research. 26 (9): 1015–35. PMID 3906008. Archived from the original on 10 اكتبر 2008. Retrieved 30 آوریل 2021. Check date values in:
|archive-date=(help) - Demopoulos CA, Pinckard RN, Hanahan DJ (October 1979). "Platelet-activating factor. Evidence for 1-O-alkyl-2-acetyl-sn-glyceryl-3-phosphorylcholine as the active component (a new class of lipid chemical mediators)". The Journal of Biological Chemistry. 254 (19): 9355–8. PMID 489536.
- Brosche T, Platt D (August 1998). "The biological significance of plasmalogens in defense against oxidative damage". Experimental Gerontology. 33 (5): 363–9. doi:10.1016/S0531-5565(98)00014-X. PMID 9762517.
- Engelmann B (February 2004). "Plasmalogens: targets for oxidants and major lipophilic antioxidants". Biochemical Society Transactions. 32 (Pt 1): 147–50. doi:10.1042/BST0320147. PMID 14748736.
- Lux H, Heise N, Klenner T, Hart D, Opperdoes FR (November 2000). "Ether--lipid (alkyl-phospholipid) metabolism and the mechanism of action of ether--lipid analogues in Leishmania". Molecular and Biochemical Parasitology. 111 (1): 1–14. doi:10.1016/S0166-6851(00)00278-4. PMID 11087912.
